DNA的复制是一个边解旋边复制的过程。复制开始时,DNA分子首先利用细胞提供的能量,在解旋酶的作用下,把两条螺旋的双链解开,这个过程叫解旋。然后,以解开的每一段母链为模板,以周围环境中的四种脱氧核苷酸为原料,按照碱基配对互补配对原则,在DNA聚合酶的作用下,各自合成与母链互补的一段子链。随着解旋过程的进行,新合成的子链也不断地延伸,同时,每条子链与其母链盘绕成双螺旋结构,从而各形成一个新的DNA分子。这样,复制结束后,一个DNA分子,通过细胞分裂分配到两个子细胞中去!
注:复制时遵循碱基互补配对原则,复制发生在细胞分裂的间期。
DNA是遗传信息的载体,故亲代DNA必须以自身分子为模板准确的复制成两个拷贝,并分配到两个子细胞中去,完成其遗传信息载体的使命。而DNA的双链结构对于维持这类遗传物质的稳定性和复制的准确性都是极为重要的。
(一)DNA的半保留复制
Waston和Click在提出DNA双螺旋结构模型时曾就DNA复制过程进行过研究,他们推测,DNA在复制过程中碱基间的氢键首先断裂,双螺旋解旋分开,每条链分别作模板合成新链,每个子代DNA的一条链来自亲代,另一条则是新合成的,故称之为半保留式复制(semiconservative replication)。 1958年Meselson和Stahl进行了如图8-3-5的实验证明了DNA分子是以半保留方式进行自我复制的。图8-3-5 Meselson和Stahl证明DNA半保留复制的实验 (二)DNA复制的起始,方向和速度
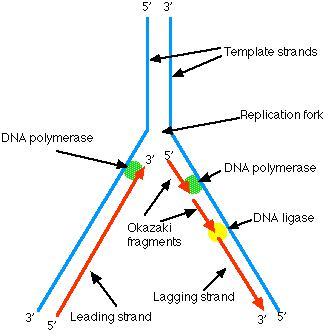
DNA在复制时,双链DNA解旋成两股分别进行。其复制过程的复制起点呈现叉子的形式,故称复制叉。以复制叉向前移动的方向为标准,一条模板链为3’—〉5’走向,在其上DNA能以5’—〉3’方向连续合成,称为前导链(leading strand);另一条模板链为5’—〉3’走向,在其上DNA也是5’—〉3’方向合成,但与复制叉移动的方向正好相反,故随着复制叉的移动形成许多不连续的冈崎片段,最后在连成一条完整的DNA链,该链称为后随链(lagging strand)。实验证明DNA的复制是由一个固定的起始点开始的。一般把生物体的单个复制单位称为复制子。一个复制子只含一个复制起点。一般说,细菌,病毒即线粒体DNA分子均作为单个复制子完成其复制,真核生物基因组可以同时在多个复制起点上进行双向复制,即它们的基因组包括多个复制子。多方面的实验结果表明,大多数生物内DNA的复制都是从固定的起始点以双向等速方式进行的。复制叉以DNA分子上某一特定顺序为起始点,向两个方向等速生长前进。图8-3-6 DNA复制过程 DNA复制过程
(三)DNA复制过程
以原核生物DNA复制过程予以简要说明
1.DNA双螺旋的解旋
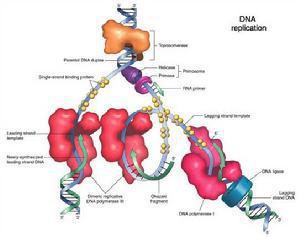
DNA在复制时,其双链首先解开,形成复制叉,而复制叉的形成则是由多种蛋白质及酶参与的较复杂的复制过程
ssbDNA蛋白是较牢固的结合在单链DNA上的蛋白质。原核生物ssbDNA蛋白与DNA结合时表现出协同效应:若第1个ssbDNA蛋白结合到DNA上去能力为1,第2个的结合能力可高达103;真核生物细胞中的ssbDNA蛋白与单链DNA结合时则不表现上述效应。ssbDNA蛋白的作用是保证解旋酶解开的单链在复制完成前能保持单链结构,它以四聚体的形式存在于复制叉处,待单链复制后才脱下来,重新循环。所以,ssbDNA蛋白只保持单链的存在,不起解旋作用。(2)DNA解链酶(DNA helicase) DNA解链酶能通过水解ATP获得能量以解开双链DNA。这种解链酶分解ATP的活性依赖于单链DNA的存在。如果双链DNA中有单链末端或切口,则DNA解链酶可以首先结合在这一部分,然后逐步向双链方向移动。复制时,大部分DNA解旋酶可沿滞后模板的5’—〉3’方向并随着复制叉的前进而移动,只有个别解旋酶(Rep蛋白)是沿着3’—〉5’方向移动的。故推测Rep蛋白和特定DNA解链酶是分别在DNA的两条母链上协同作用以解开双链DNA。(3)DNA解链过程 DNA在复制前不仅是双螺旋而且处于超螺旋状态,而超螺旋状态的存在是解链前的必须结构状态,参与解链的除解链酶外还有一些特定蛋白质,如大肠杆菌中的Dna蛋白等。一旦DNA局部双链解开,就必须有ssbDNA蛋白以稳定解开的单链,保证此局部不会恢复成双链。两条单链DNA复制的引发过程有所差异,但是不论是前导链还是后随链,都需要一段RNA引物用于开始子链DNA的合成。因此前导链与后随链的差别在于前者从复制起始点开始按5’—3’持续的合成下去,不形成冈崎片段,后者则随着复制叉的出现,不断合成长约2—3kb的冈崎片段。
2.冈崎片段与半不连续复制
因DNA的两条链是反向平行的,故在复制叉附近解开的DNA链,一条是5’—〉3’方向,另一条是3’—〉5’方向,两个模板极性不同。所有已知DNA聚合酶合成方向均是5’—〉3’方向,不是3’—〉5’方向,因而无法解释DNA的两条链同时进行复制的问题。为解释DNA两条链各自模板合成子链等速复制现象,日本学者冈崎(Okazaki)等人提出了DNA的半连续复制(semidiscontinuous replication)模型。1968年冈崎用3H脱氧胸苷短时间标记大肠杆菌,提取DNA,变性后用超离心方法得到了许多3H标记的,被后人称作冈崎片段的DNA。延长标记时间后,冈崎片段可转变为成熟DNA链,因此这些片段必然是复制过程中的中间产物。另一个实验也证明DNA复制过程中首先合成较小的片段,即用DNA连接酶温度敏感突变株进行试验,在连接酶不起作用的温度下,便有大量小DNA片段积累,表明DNA复制过程中至少有一条链首先合成较短的片段,然后再由连接酶链成大分子DNA。一般说,原核生物的冈崎片段比真核生物的长。深入研究还证明,前导链的连续复制和滞后链的不连续复制在生物界具有普遍性,故称为DNA双螺旋的半不连续复制。
3.复制的引发和终止
所有的DNA的复制都是从一个固定的起始点开始的,而DNA聚合酶只能延长已存在的DNA链,不能从头合成DNA链,新DNA的复制是如何形成的?经大量实验研究证明,DNA复制时,往往先由RNA聚合酶在DNA模板上合成一段RNA引物,再由聚合酶从RNA引物3’端开始合成新的DNA链。对于前导链来说,这一引发过程比较简单,只要有一段RNA引物,DNA聚合酶就能以此为起点,一直合成下去。对于后随链,引发过程较为复杂,需要多种蛋白质和酶参与。后随链的引发过程由引发体来完成。引发体由6种蛋白质构成,预引体或引体前体把这6种蛋白质结合在一起并和引发酶或引物过程酶进一步组装形成引发体。引发体似火车头一样在后随链分叉的方向前进,并在模板上断断续续的引发生成滞后链的引物RNA短链,再由DNA聚合酶 III 作用合成DNA,直至遇到下一个引物或冈崎片段为止。由RNA酶H降解RNA引物并由DNA聚合酶 I 将缺口补齐,再由DNA连接酶将每两个冈崎片段连在一起形成大分子DNA.。 (四)端粒和端粒酶
1941年美籍印度人麦克林托克(Mc Clintock)就提出了端粒(telomere)的假说,认为染色体末端必然存在一种特殊结构——端粒。已知染色体端粒的作用至少有二:① 保护染色体末端免受损伤,使染色体保持稳定;② 与核纤层相连,使染色体得以定位。 在弄清楚DNA复制过程之后,20世纪70年代科学家对DNA复制时新链5’端的RNA引物被切除后,空缺是如何被填补的提出了质疑。如不填补岂不是DNA每复制一次就短一点。以后随链复制为例,当RNA引物被切除后,冈崎片段之间是由DNA聚合酶 I催化合成的DNA填补之,然后再由DNA连接酶将它们连接成一条完整的链。但是DNA聚合酶 I 催化合成DNA时,需要自由3’—OH作为引物,最后余下子链的5’无法填补,于是染色体就短了一点。 在正常体细胞中普遍存在着染色体酶复制一次端粒就短一次的现象。人们推测,可能一旦端粒缩短到某一阈限长度一下时,他们就会发出一个警报,指令细胞进入衰老;或许是当细胞判断出它们的染色体已变得太短了,于是分裂也就停止了,造成正常体细胞寿命有一定界限。但是在癌细胞中染色体端粒却一直维持在一定长度上,这是为什么?这是因为DNA复制后,把染色体末端短缺部分补上需要端粒酶,这是一种含有RNA的酶,它既解决了模板,又解决了引物的问题。在生殖细胞和85%癌细胞中都测出了端粒酶具有活性,但是在正常体细胞中却无活性,20世纪90年代中期,Blackburn首次在原生动物中克隆出端粒酶基因。
端粒酶在癌细胞中具有活性,它不仅使癌细胞可以不断分裂增生,而且它为癌变前的细胞或已经是癌性的细胞提供了时间,以积累附加的突变,即等于增加它们复制,侵入和最终转移的能力。同时人们也由此萌生了开发以端粒为靶的药物,即通过抑制癌细胞中端粒酶活性而达到治疗癌症的目的。
至于真核细胞DNA末端的结构特点,早就在1978年Blackburn就以原生动物四膜虫(一种纤毛虫)为例说明之:① 迥纹形式的发夹环;② 仅由C,A组成的简单序列大量重复(C4A2)20~70;③ 链上有许多缺口(nicks)。