
精选百科
本文由作者推荐
基本信息
| 中文名 | 内含子 |
| 外文名 | Intron |
| 界 | 真菌界 |
| 门 | 真核结构门 |
| 纲 | 细胞纲 |
| 科 | 细胞科 |
| 定义 | 一个基因或mRNA分子中无编码作用的片段 |
| 别称 | 间隔顺序 |
收起
定义
内含子
内含子(introns)在转录后的加工中,从最初的转录产物除去的内部的核苷酸序列。术语内含子也指编码相应RNA内含子的DNA中的区域。内含子可能含有“旧码”,就是在进化过程中丧失功能的基因部分。正因为内含子对翻译产物的结构无意义,它比外显子累积有更多的突变。
特点
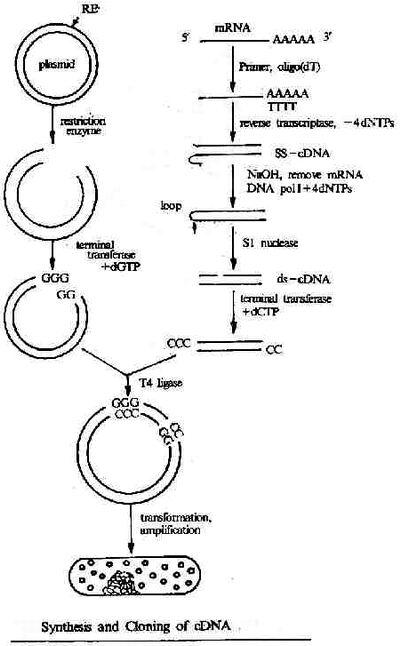
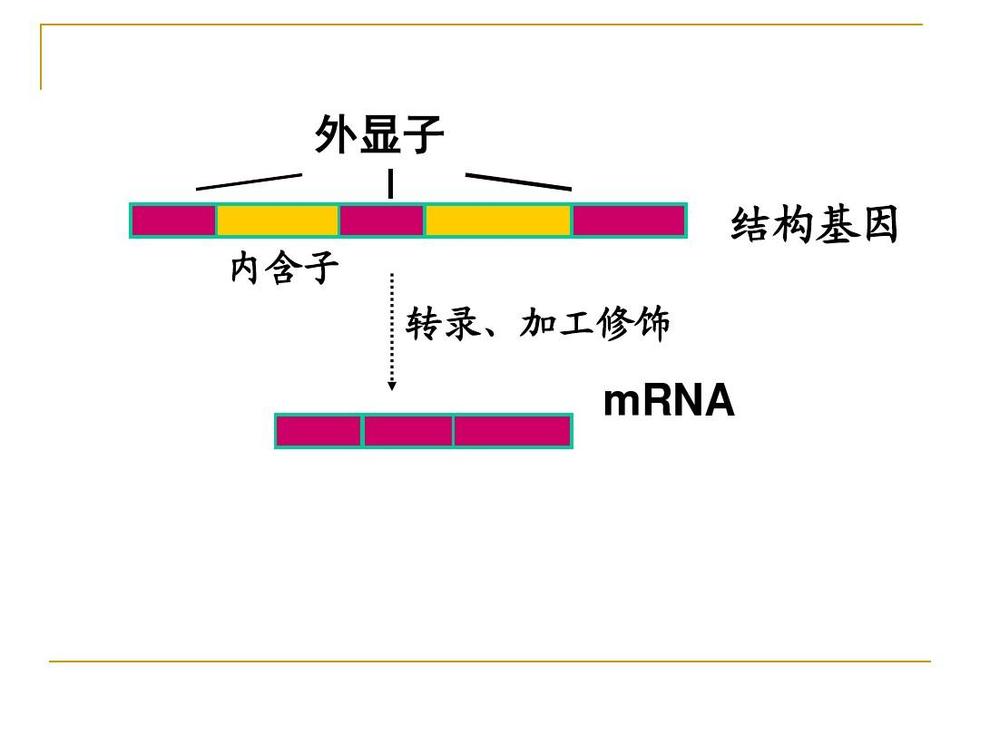
大多数真核结构基因中的间插序列(interveningsequence)或不编码序列。它们可以转录,但在基因转录后,由这些间插序列转录的部分(也可用内含子这个术语表示)经加工被从初级转录本中准确除去,才产生有功能的RNA。基因的编码部分称外显子。内含子常比外显子长,且占基因的更大比例。真核基因所含内含子的数目、位置和长度不尽相同,如鸡卵清蛋白基因的外显子被7个内含子隔开,鸡卵伴清蛋白基因有17个内含子,α-珠蛋白基因有2个内含子,卵粘蛋白基因有6个内含子等。
内含子(Interveningregion)是一个基因中非编码DNA片断,它分开相邻的外显子。更精确的定义是:内含子是阻断基因线性表达的序列。DNA上的内含子会被转录到前体RNA中,但RNA上的内含子会在RNA离开细胞核进行转译前被剪除。在成熟mRNA被保留下来的基因部分被称为外显子。真核生物的基因含有外显子和内含子,是前者区别原核生物的特征之一。
内含子和外显子的比例因种而异。河鲀鱼的内含子比较少。
但内含子与垃圾DNA不同,垃圾DNA亦即那些基因以外的序列,还未被发现有任何功能的DNA,但可能是参与基因调控和选择性剪接的调控。但若内含子对应的mRNA片断若没有被除去,可能会发生非常大的突变。如一种植物,科学家抑制了剪切酶的活性而保留了其mRNA中一段内含子。结果,该植物的雌蕊发育不正常。而雄蕊却出现了雌蕊的特征。
“马赛克基因”,就是说编码的DNA片断(外显子)被非编码区域(内含子)隔开,该概念是1977年由Hogness,Mandel和Chambon提出。
信息
相位
类型
第一类内含子
内含子
一、第一类内含子可动现象的发现和被步研究
1975年,人们以啤酒酵母菌mt-DNA某些突变为标记进W+XW-杂交,发现W+传递到了代的比例为95%而W-几乎为零,现象上好似发生了单方赂基因转变(unidirectionalgeneconversion)。从W-到W+,由于W+中有内含子ScLDU.1,而W-则无,故人们认为上述现象与该内含了有关。顺序分析发现该内含子中存在一个可读框PRF,起始于AUG,共长235个密码子,为确定该产物的生化功能,人们以内含子上下游的一些富含GC的顺序作为探针来研究W+XW-杂交中两不同阶段的mtDNA,即:(1)合子刚形成时的mt-DNA,此时双亲的mt-DNA同时存在于一个细胞中;(2)几个细胞世代以后,当细胞是同质体时的mt-DNA。通过对W+XW-的子代中的同质体克隆的观察发现,内含子ScLSU.在子代中的传递效率高达95%,恰好与W+情况相吻合,而mt-DNA上远离ScLSU.内含子发生了单方向基因转变,而且在单方向基因转变同时还发生距该内含子上下游几百bp范围的侧翼DNA顺序的共转变,其频率随距内含子的距离的增加而减小。
而当研究W+XW-中的合子刚形成时的mt-DNA时,发现在无内含子(intronless)LSU基因的内含子插入位发生一个双链断裂,该断裂将随合子的长大而消失,推测与内含子插入引起的DNA修复有关。同时,当Wd突变体和无内含子的野生型杂交时,发现没有内含子的单方向基因转变发生,核苷酸顺序分析表明Wd突变原因是在内含子ORF的不同位置发生了移码突变、无义突变和多个碱基置换,说明该内含子的翻译产物在I-(intronless)基因的双链断裂形成中起着直接或间接的作用,1986所有人用通用遗传密码构建了一个等同于ScLSU.1内含子ORF的顺序,发现它在E.coli中能表达为一个内切核酸酶,并可高度特异地切割带内含子切割序的mt-DNA,该识别顺序共长18bp,切割后可产生4bp伸出的3'-OH末端以插入内含子,而内含子一旦插入到无内含子顺序后,该顺序就被分割成两部分,无法再为内切核酸酶识别,从而可阴止I+基因的自切割,使基因变均表现为单方向的,即从I+到I-。通过对酵母W系统的研究可概括出第一类内含子可动的三个条件:
(1)一个基因的两种功能形式的存在,即I+和I-,且能通过杂交进行遗传交换;
(2)在内含子中要存在一咱ORF,并编码有功能的双链内切核酸酶;
(3)在无内含子基因中存在呆为该内切梳酸酶识别的顺序。
在W系统研究之后,人们通过许多杂交实验又发现了一系列可动的第一类内含子,例Chlamydomonaseugametos的cp-DNA中的CeLSU.5、Saccharomycescererisiae中的Sccox1.4,T4噬菌体中的T4td.1和T4aunY.1,Physarumpolycephalum的PpLSU.3及Neurosporacrassa中的含子Ncnd1,1等。
内含子和 外显子
第一类内含了DNA的内部ORF表达,产生一个双链内切核酸酶,该酶识别无内含子的同一基因上的顺序并结合上去,酶切产生具有4bp伸出析3'-OH末端的双链断裂,然后以I+基因的未切割的同源顺序作为模板来修复双链断裂,该修复机制延伸到侧翼区域,导致共转变,而内含子一旦插入后,它本身既可产生内切核酸酶,也可作为修复断裂的模板,从而使整个过程可不断进行。
内含子转座也通过相同的过程进行,由于内切核酸酶识别顺序可存在于三种位置:(1)同一基因的拷则;(2)不同基因;(3)基因间区域,这三种情况下的内含子插入,导致三种不同结果,第一种情况在内含子插入后产生内含子回归;第二种导致转座,因为插入新位点的内含子进行有效的RNA剪接;第三种情况也导致转座,但由于插入的内含子DNA无法转录,故它将经随机突变而消失。同时,在双链断裂的修复中,切点周围的顺序和未切割模板的外显顺序之间的同源性对修复亦很重要,同源性越高,修复效率越高。所以,内含子回归很常见,而内含子转座则很少见。
另处,内含子转座也可通过RNA中介进行,基因Gl(I+)经转录和自剪接产生被切下来的内含子RNA,该RNA通过反向剪接整合到另一个基因G2(I-)中,再经逆转录和重组,产生带有I的G2,这种情况主要是在四膜虫中发现的。
内含子
内含子DNA顺序插入到一个新位置,应带来它的转录和剪接问题,内含子回归不存在这个问题,因为I+基因本身可正常转录和剪接。但是,就内含子转坐而言,由于内含子本身无启动子,其转坐有赖于宿主基因,所以,内含子转座常导致产生一个无活性的内含子,它将因无法时行有效转录而通过随机突变在进化中消失,故内含子转座的频率很低。
同时,当内含子插入到某一基因后,要保持该基因的正常功能,就必须有效的进行RNA剪接,第一类内含子的剪接和内含子的IGS(internalguidesequence)顺序与上游外显子之间碱基配对的相互作用有关,故一个成功的内含子插入依赖于该内含子能够与新的基因相匹配。同时,资料表明,许多第一类内含子的上游外显子有短的保留下游外显子没有,故推测可能内含子发生转座后,可通过随机突变或特异的复制机来适应新的回归位点,以获得与外显子的正确匹配,从而成为一个成功的内含子转座,当然,转座中外显子的共转变或外显子通过随机突变而适应新插入的内含子两种方式同样也能提高内含子和外显子的匹配,提高内含子转座的成功率。
既然单方向内含子插入的频率及高,I-等位基因为何未在群体中消失呢?一种解释是:I+基因是近期出现的,并将随时间推移取代所有I-等位基因;另一种解释认为是自然不利于含有内含子的细胞的生长;而目前更多的人认为是内含子回归和丢失之间的平衡维持第一类内含在细胞世代稳定存在,只要插入到合适的位置后能进行有效的转录和剪接,同时不给细胞带来任何新的表型。在S.cerevisiae中,mt-内含子的剪接功能缺陷型是呼吸缺陷的,而这些内含子发生缺失后就可得到呼吸正常的野生型表型的回复子,故人们推测内含子丢失的可能机制是:I+RNA发生剪接,除去内含子,再反转录,并通过受体基因和cDNA外显子顺序之间的同源重组,导致内含子丢失,而实验中发现第二类内含子和反转录病毒的逆转录酶有同源顺序,
使人们猜想第一类内含子的丢失可能由第二类内含子的翻译产物控制,1989年有人用mss18突变体证明了这一想法,因为实验表明,第一类内含子Secoxl.5b的发生缺失需要第二类内含子Sccoxl.1Sccoxl.2的存在。
目前对大量的生物研究表明,大批内含子在生物中的分布物不均衡,表示为(1)同一种的不同个体中,有的有内含子,有的没有;(2)同一基因在不同种生物基因组中的内含子的特性,数目位置等不同,如Sccoxl.2b和Ancoxl.3是高度同源内含子插入在不同种的同一回归位点的例子,这两种内含子有70%顺序相同,但内含子周围的外显子顺序很不同,从而推测可能是内含子发生水平转移的结果,而不同生物不同回归位点有高度同源内含子存在的事实,如ScLSU.1和Nccob.2也说明水平转移的可能。总之,内含子在不同种或同一种的不同个体中的不同回归位点分布的不均可以用内含子回归和内含子丢失之间的平衡来解释,而在不回归位点分布的不均可以用回归的内含子丢失之间的平衡来解释,而在不同回归位点上相似相关内含子的存在则可能是内含子转座的结果。而且,如果这种内含子转座是发生在极其不同的种之间的话,那么,可能在转移过程中还存在着某种未知形式的分子载体。当然,这还只是一种设想,有待于进一步的证实。
起源
内含子起源有两种假说。
1.内含子与它所在的基因一样古老,在装配第一个这样的基因时,内含子就已存在。早期的内含子具有自催化、自我复制等能力,因此,它们是原始基因和基因组的组织与复制必不可少的部分。而今天的原核生物和少数低等的真核生物,由于它们需要进行快速的DNA复制从而进行快速的细胞分裂,因而失去了内含子。现代的内含子是一类进化遗迹,它们之所以能继续存在,是因为具有重新组合基因组中的外显子以形成新的基因的能力,即内含子能赋予其携带者更大的进化潜力。
2.内含子不是基因原有的,而是在进化的某一过程中通过转座作用插入到连续基因中去的,内含子在较高级的功能基因或在真核生物出现之后才产生。这种假说必须面对一个难题,即内含子最初如何能插入到连续编码的基因中而保持基因的功能不变。
内含子相关的文章

小编整理:神功是武则天使用的年号,这个年号属于唐朝,使用时间是679年九月—681年二月。此外,这个年号是武则天在位期间使用的最后一个年号,她在位期间还使用了其他一些年号,比如光宅、垂拱、永昌、载初等。总的来说,神功是一个历史上的年号,它代表了一个时期的历史事
巴尔干半岛(Balkan Peninsula),是一个历史和地理上的名词,用以描述欧洲的东南隅位于亚得里亚海和黑海之间的陆地,详细的范围依照定义不同有许多种说法。巴尔干一词是由土耳其语的“山脉”一词派生出来的。
胡杨(拉丁学名:Populus euphratica),别名:胡桐、英雄树、异叶胡杨、异叶杨、水桐、三叶树、幼发拉底杨等,是杨柳科(Salicaceae)杨属(Populus)一种落叶中型天然乔木植物。主要分布于蒙古、埃及、叙利亚、印度、伊朗、阿富汗、巴基斯坦等地区,在中国主要分布于内蒙古西部、甘肃
西晋(公元265年—公元316年)是中国历史上三国时期之后的大一统王朝之一,另与东晋合称晋朝。传四帝,国祚五十一年。若以灭东吴始,则仅立朝37年。为了区别于东晋,史称西晋,两晋又被称为司马晋。
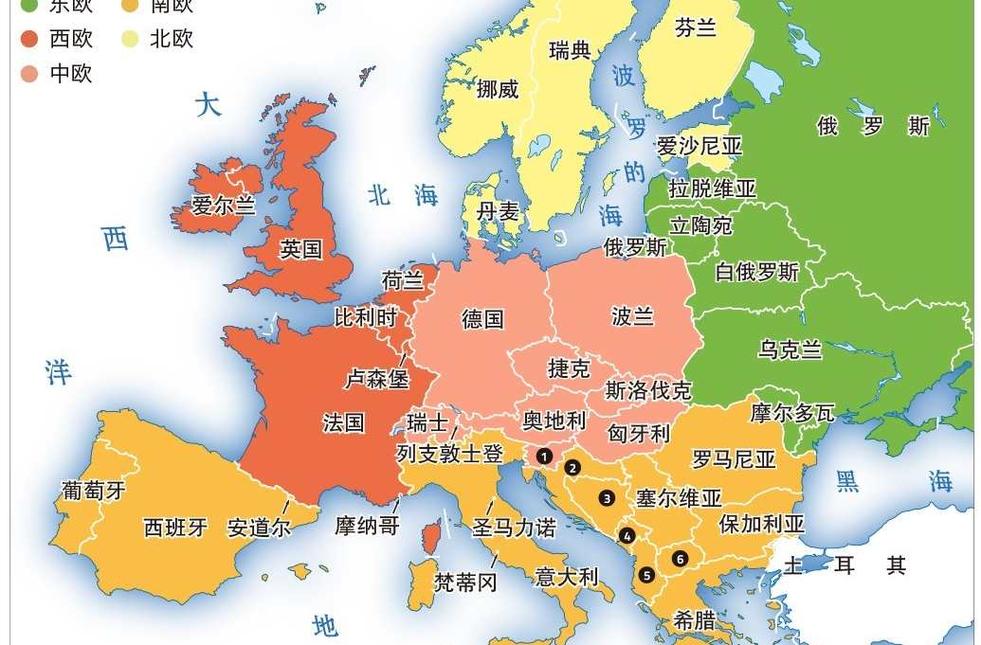
南欧欧洲南部的简称南欧(southern Europe)是欧洲南部的简称,范围包括伊比利亚半岛、亚平宁半岛及巴尔干半岛南部,包括西班牙、葡萄牙、安道尔、意大利、希腊、马耳他、梵蒂冈、圣马力诺、斯洛文尼亚、克罗地亚、阿尔巴尼亚、罗马尼亚、保加利亚、塞尔维亚、黑山、马其顿和波斯尼亚和黑塞哥维那共17个国家,也称为地中海欧洲,因为大多南欧国家靠近地中海。面积约为166万多平方公里。南欧隔着地中海与亚、非

尚可名片
这家伙太懒了,什么都没写!
作者
