
旱獭(学名:Marmota bobak):是松鼠科中体型最大的一种,常称为土拨鼠,含3个亚种。大型啮齿动物,体长为500毫米,体重4-5千克。尾短为110毫米。在外形和生活方式上都与鼠类相似,体型粗大肥壮,颈粗吻阔,耳小眼细,四肢粗短,利爪坚硬,松尾短扁。
栖息于草原、低山丘陵区。是陆生和穴居的草食性动物,主要食物为草、浆果、地衣、苔藓、根和花。冬眠。因潜藏烈性病菌,又危害牧场,一向被疾控与植保部门列入监控、杀灭黑名单。分布于哈萨克斯坦、俄罗斯联邦和乌克兰。哺乳动物无危陆生动物杂食性

旱獭(学名:Marmota bobak):是松鼠科中体型最大的一种,常称为土拨鼠,含3个亚种。大型啮齿动物,体长为500毫米,体重4-5千克。尾短为110毫米。在外形和生活方式上都与鼠类相似,体型粗大肥壮,颈粗吻阔,耳小眼细,四肢粗短,利爪坚硬,松尾短扁。
栖息于草原、低山丘陵区。是陆生和穴居的草食性动物,主要食物为草、浆果、地衣、苔藓、根和花。冬眠。因潜藏烈性病菌,又危害牧场,一向被疾控与植保部门列入监控、杀灭黑名单。分布于哈萨克斯坦、俄罗斯联邦和乌克兰。哺乳动物无危陆生动物杂食性
旱獭
土拨鼠
约7~10千克
脊椎动物亚门
松鼠科
动物界
啮齿目
脊索动物门
哺乳纲
地松鼠族
旱獭
东欧和中亚
Marmota bobak
hàn tǎ
无危(LC) IUCN标准
兽亚纲
松鼠形亚目
非洲地松鼠亚科
西部亚种、东部亚种
Muller、1776
杂食性
陆生动物
无危
旱獭属
约490~575毫米
Bobak Marmot
旱獭是大型啮齿动物。体短身粗,成年长490-575毫米,体重7-10千克。无颈,尾、耳皆短,耳壳黑色。头骨粗壮,长度89-103毫米。上唇为豁唇,上下各有一对门齿露于唇外,两眼为圆形,眶间部宽而低平,眶上突发达,骨脊高起,有更宽颧弓,头骨身体各部肌腱发达有力。四肢短而强,前足4趾,后足5趾,可直立行走;母獭有6-7对乳头。前爪发达,适于掘土。
被毛通常短而密,粗糙,毛色因地区、季节和年龄变异。春季毛色淡。背部毛呈黄褐或淡褐色。腹部为土黄色,毛基色褐灰,体侧和腹侧的毛色无显著分界线,头部及尾部色较深。各不同种旱獭毛色深浅略有差异。

旱獭
物种对比对比维度 | 旱獭 | 土拨鼠 | 水獭 | 树懒 |
| 外观 |
|
|
|
|
| 体长 | 约490~575毫米 | 体长37~40厘米 | 70~75厘米 | - |
| 体重 | 约7~10千克 | 4~8千克 | - | - |
| 分布区域 | 东欧和中亚 | 分布范围仅限于南亚西北部和中国的高海拔地区,其遍布于印度、尼泊尔和巴基斯坦的喜马拉雅山脉以及中国的西部、中部和南部的青藏高原地区 | 四川省、福建省、广东省、湖南省、浙江省等地 | 中美洲、南美洲 |
| 食性 | 杂食性 | 植食性 | 肉食性 | 植食性 |
旱獭主要栖息于气候寒冷的丘陵地区、山地的各种草原和高山草甸,温带草原和半荒漠地区。[1]
分布于哈萨克斯坦、俄罗斯联邦和乌克兰。
20世纪初,该物种分布在从乌克兰西部到俄罗斯和哈萨克斯坦到额尔齐斯河的整个草原地带。然而,在20世纪上半叶,由于狩猎和栖息地丧失,大草原的耕作和耕地的转变,从而大大减少了欧洲大草原旱獭的分布范围。到1940年代,旱獭在欧洲范围已在未使用的土地和保护区内分裂成孤立的种群。分布范围的大部分限于乌拉尔和哈萨克斯坦北部。
集群穴居,挖掘能力甚强,洞道深而复杂,多挖在岩石坡和沟谷灌丛下。从洞中推出的大量沙石堆在洞口附近,形成旱獭丘。白天活动,食量大,每日啃食大量优良牧草,最喜欢的植物有野燕麦、冰草、菊苣、三叶草、和小旋花(田旋花)。偶尔也吃菜园草、向日葵和农作物,如土豆。耐饥饿,不饮水,喜食含水量大的多汁饲料。

旱獭
野栖旱獭主要以莎草科、禾本科植物的叶、茎,豆科植物的花为食,且有季节性变化,在饲养条件下表现为杂食性。旱獭易驯化,不伤人,不耐热,怕曝晒,抗病力强。当气温长时间低于10℃以下时,就自然冬眠,时间可长达3-6个月,当气温转暖后自然苏醒。
穴居
有家族性的同居生活,通常一个家族占据由各种类型的洞穴组成的洞群,称为家族洞群。由邻近数个家族共同组成一个群聚。家族之间的个体相互之间和睦相处。一个群聚中的个体活动小区及取食领地,可以互相重叠。个体之间的接触是相当频繁与密切的,彼此之间常常有互相迁入或迁出,重组家族的现象时常发生。至于家族洞群间所分布的一些临时洞,则更是成为各个家族成员暂时息小憩的共用场所。一般在不同季节挖掘不同的洞穴,有主洞、副洞、临时洞等多种型。主洞又叫冬眠洞,主要作为冬眠之用,但也可以作为养育幼仔的洞巢,洞道的结构极为复杂,就像一个地下宫殿,工程十分浩大,需要搬出数量惊人的土石,洞深可达数米之长,洞道的分支也较多,总长度可以达到十多米,在洞道的深处还有一个椭圆形的巢室,巢室内铺垫着干草,而且要定期清理、更换和变动位置,一般都有好几个出入的洞口,洞口十分光滑,并有一股特殊的骚味,常招引很多蝇类和其他昆虫营集,附近还常常设有厕所,其中往往堆积着大量的粪便。冬眠的时候在洞中存有大量食物并铺有干草,里面用干草、湿泥和粪便等将洞口堵住。副洞的洞道比较浅,结构也较为简单,分支极少,或仅仅只有一个单一的洞道,出入口一般也只有一个,多为雄性成年个体夏季的居住之所。临时洞构造更是极为简单,洞道极浅,仅仅作为临时休息或应急避敌之用。同一个家族的各个洞口之间都有通道相连接,甚至相邻家族的洞群之间,也常有小通道连接在一起。此外,在它栖息的范围内,还常常分布着一些废弃的洞穴,这是由于死亡或迁出之后遗留下来的,洞口有时杂草丛生,结有蛛网或塌陷,但不久就会有附近的其他个体迁入,或者被其他小型啮齿动物和野兔等占用。
活动
属于白昼活动的动物,尤以早晨和黄昏最为活跃。早上出洞的时间随季节而异,一般依太阳照射到洞口来确定。每次出洞之前总是先探出头来四处张望,觉得安全后,先露出半个身子,扒在洞口晒晒太阳,然后发出鸣叫声。此时,临近的同类立即响应,一起鸣叫。此后不久,即开始取食,除非是遇有敌害外,则在这以后的一天内完全不再发声鸣叫。日落之前进入洞中休息,夜间不再出来活动。
食物
主要以草本植物为食,喜欢吃带有露水珠的嫩草茎叶、嫩枝或草根,尤其是莎草科、禾本科和豆科植物的地上绿色的部分,偶尔也取食一些昆虫和小型啮齿动物。在农作区,它也常常偷食青稞、燕麦、油菜、洋芋等作物的禾苗、茎叶。早春,在青草尚未发芽时,则也可挖食草根。
冬眠
集群穴居,秋季体内积存大量脂肪,秋后闭洞处蛰眠状态,次年春季3-4月份出洞活动。
旱獭的寿命可长达15-20年,繁殖年限为10-15年。春季是旱獭配种的季节,出蛰后十天左右开始交配。一般年产1次,雌獭妊娠期为40-42天,每胎产仔4-6只,多者达12只以上。初生体重约27克,36-40月龄性成熟。其种群繁殖力与种群密度有关,当种群密度大幅下降时,繁殖大幅提高。
出蛰后不久即交配繁殖。幼獭于第3年性成熟。一年繁殖一次,出蛰后不久即进入繁殖期,开始交配,延续约1个月左右,个体活动极其频繁,经常串洞、追逐,以进行性活动为主,吃食时间很短,很少警戒,活动范围很大,其中尤以成年雄兽参与繁殖的个体的活动性最强。4月中旬即可发现怀孕的雌兽,怀孕期约为35天左右,每胎产1-9仔,而以4-6只为最多见。幼仔出生后,雌兽吃食时间与范围逐渐增加,为保护幼兽守望警戒增多,串洞和交往则显减少。6月底即可见到幼仔出洞活动,十分活跃,取食频繁。幼体与母兽一直生活至第2年的7月才分居出去,独立生活。3岁时达到性成熟。但每年参与繁殖的雌性个体,仅仅只占达性成熟雌性个体总数的50-60%。
场地
旱獭的建造场地必须是地热干燥、平坦、背风遮阳,且少干扰的僻静地方,因此,应远离其他饲养场、远离屠宰场、畜产品加工厂,要单独饲养,不要与其他动物混养。
建窖
旱獭饲养方法常见有舍饲方式、棚饲方式、地窖方式以及地沟方式等。其中以地窖方式简便而近于旱獭的自然状态,较适合家庭饲养使用。窖址选择宽敞地形,避开路边、作坊等僻静处所,土质应坚实,地下水位较低处作为饲养的窖址。
饲料
饲料主要是青饲料如新鲜牧草、蔬菜等精饲料以玉米、麦麸等。
| 旱獭(3亚种) | |||
| 中文名称 | 学名 | 命名者及年代 | |
| 1 | 旱獭指名亚种 | Marmota bobak bobak | Müller, 1776 |
| 2 | 旱獭伏尔加河亚种 | Marmota bobak kozlovi | Fokanov,1966 |
| 3 | 旱獭哈萨克斯坦亚种 | Marmota bobak tschaganensis | Bazhanov, 1930 |
| 旱獭属(14种) | |||
中文名称 | 学名 | 英文名称 | |
1 | 灰旱獭 | Marmotabaibacina | AltaiMarmot |
2 | 旱獭 | Marmotabobak | BobakMarmot |
3 | 阿拉斯加旱獭 | Marmotabroweri | AlaskaMarmot |
4 | 蒙塔古岛褐旱獭 | Marmotacaligata | MontagueIslandHoaryMarmot |
全部列入《世界自然保护联盟》(IUCN)2016年濒危物种红色名录ver3.1。
种群现状
以前,旱獭在从乌克兰西部到额尔齐斯河的整个草原地带都很常见和丰富。到1940年代,欧洲数量和分布范围急剧下降,该物种仅限于未开垦地区和自然保护区的孤立碎片栖息地。在1960年代,俄罗斯和乌克兰都禁止狩猎,这些国家的旱獭数量随后增加,已经被认为是稳定的。相比之下,在哈萨克斯坦北部,数量尚未恢复并保持低密度,但在哈萨克斯坦中部,数量正在增加。该物种在1980年代初被重新引入许多地方,该物种也自然地重新定居在许多地区。
旱獭全身均是宝:其肉可食;脂肪可入药,内可治咯血,外可治烧伤,还可加工制成高级化妆品,具有润肤、护肤作用。旱獭毛皮的皮质较好,坚实耐磨,旱獭绒毛的色染性能较佳,加工后的毛色光亮鲜艳,用以制成裘时,工艺价值很高。
1.Marmots of the World · marmotburrow.ucla(引用日期:2016-03-31)
2.Marmota Blumenbach, 1779 · BioLib.cz(引用日期:2016-03-31)
3.Marmota · The IUCN Red List(引用日期:2017-12-03)

缺血性脑血管病是指颅内外动脉狭窄或闭塞导致的脑血管病,是不同程度的缺血性脑血管疾病的总称。包括短暂性脑缺血发作、脑梗死。所有影响脑血管壁的结构和功能、血液成分及血流动力学的各种因素,都可以成为缺血性脑血管病的病因。临床表现有视力减退或失明、感觉和运动障碍、偏瘫、意识障碍、昏迷、失语、饮水呛咳、吞咽困
黄蓉是《射雕英雄传》女主角、《神雕侠侣》重要配角,在《倚天屠龙记》中,也曾引述过和黄蓉相关的事迹。她是“东邪”黄药师独女、“北丐”洪七公徒弟、丐帮第十九代帮主、南宋大侠郭靖的妻子,娇美无比,风姿绰约,武功高强,擅长打狗棒法、玉箫剑法、《九阴真经》功法、落英神剑掌、阴阳五行和八卦术数等
沙特阿拉伯王国(阿拉伯语:مملكةعربيةسعودية,英语:Kingdom of Saudi Arabia),通称沙特阿拉伯(Saudi Arabia),简称沙特。沙特位于阿拉伯半岛。东濒波斯湾,西临红海,同约旦、伊拉克、科威特、阿联酋、阿曼、也门等国接壤,并经法赫德国王大桥与巴林相接。海岸线
广东,简称“粤”,省会广州,中华人民共和国省级行政区。因古地名广信之东,故名“广东”。位于南岭以南,南海之滨,与香港、澳门、广西、湖南、江西及福建接壤,与海南隔海相望。受地壳运动、岩性、褶皱和断裂构造以及外力作用的综合影响,广东省地貌类型复杂多样,有山地、丘陵、台地和平原等。广东是中国的南大门,处在
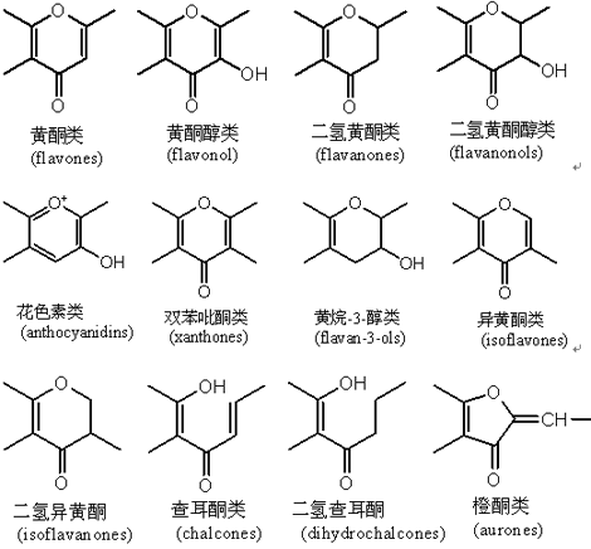
黄酮黄酮类化合物的总称黄酮(flavone),是黄酮类化合物的总称,泛指两个具有酚羟基的苯环(A-与B-环)通过中央三碳原子相互连结而成的一系列化合物。黄酮类化合物结构中常连接有酚羟基、甲氧基、甲基、异戊烯基等官能团。黄酮类化合物(英语:Flavonoid,又称类黄酮)是指基本母核为2-苯基色原酮类化合物,现在则泛指两个具有酚羟基的苯环通过中央三碳原子相互连接的一系列化合物。他们来自于水果、蔬菜、

尚可名片
这家伙太懒了,什么都没写!






