
操纵子学说是1961年,法国科学家莫诺(J·L·Monod,1910-1976)与雅可布(F·Jacob)发表“蛋白质合成中的遗传调节机制”一文中提出的学说,该学说开创了基因调控的研究。
精选百科
本文由作者推荐
操纵子学说
操纵子学说
中文名
操纵子学说
提出者
莫诺(J·L·Monod,1910-1976)与雅可布(F·Jacob)
提出时间
1961年
应用学科
医学
调节内容
基因编码调节蛋白
适用领域
医学
介绍

色氨酸操纵子
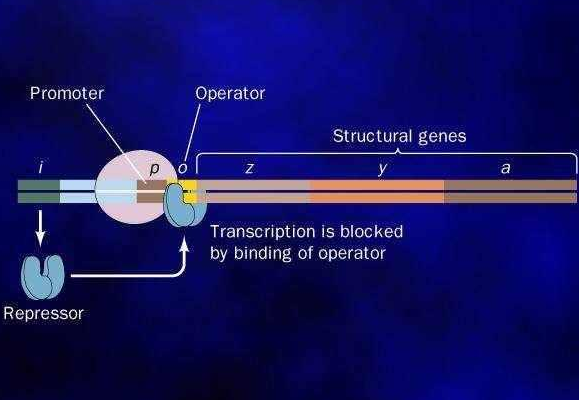
1961年,法国科学家莫诺(J·L·Monod,1910-1976)与雅可布(F·Jacob)发表“蛋白质合成中的遗传调节机制”一文,提出操纵子学说,开创了基因调控的研究。四年后的1965年,莫诺与雅可布即荣获诺贝尔生理学与医学奖。莫诺与雅可布最初发现的是大肠杆菌的乳糖操纵子。这是一个十分巧妙的自动控制系统,这个自动控制系统负责调控大肠杆菌的乳糖代谢。
乳糖可作为培养大肠杆菌的能源。大肠杆菌能产生一种酶(叫做“半乳糖苷酶”),能够催化乳糖分解为半乳糖和葡萄糖,以便作进一步的代谢利用。编码半乳糖苷酶的基因(简称z)是一个结构基因(structural gene)。这个结构基因与操纵基因共同组成操纵子。操纵基因受一种叫作阻遏蛋白的蛋白质的调控。当阻遏蛋白结合到操纵基因之上时,乳糖会起诱导作用,它与阻遏蛋白结合,使之从操纵基因上脱落下来。这时,操纵基因开启,相邻的结构基因也表现活性,细菌就能分解并利用乳糖了,这样,乳糖便成了诱导半乳糖苷酶产生的诱导物。
上述内容表明,大肠杆菌的乳糖操纵子是一个十分巧妙的自动控制系统:当培养基中含有充分的乳糖,同时不含葡萄糖时,细菌便会自动产生半乳糖苷酶来分解乳糖,以资利用。当培养基中不含乳糖时,细菌便自动关闭乳糖操纵子,以免浪费物质和能量。
启动基因
60年代中期,在操纵子中还发现了另一个开关基因,称为启动基因(promoter)。启动基因位于操纵基因之前,二者紧密相邻。启动基因由环腺苷酸(cAMP)启动,而环腺苷酸能被葡萄糖所抑制。这样,葡萄糖便通过抑制环腺苷酸而间接抑制启动基因,使结构基因失活,停止合成半乳糖苷酶。
由此可知,结构基因同时受两个开关基因——操纵基因与启动基因的调控。只有当这两个开关都处于开启状态时,结构基因才能活化。当培养基中同时存在葡萄糖和乳糖时,葡萄糖通过抑制环腺苷酸而间接抑制启动基因,并进而抑制结构基因,使细菌不产生半乳糖苷酶。这种情况下,细菌便会自动优先利用葡萄糖,因为葡萄糖果是比乳糖更好的能源。
1969年,贝克维斯(J·R·Beckwith)从大肠杆菌的DNA中分离出乳糖操纵子,完全证实了雅可布和莫诺的模型。
在启动基因发现之前,莫诺和雅可布的操纵子模型中,直接对结构基因起操纵作用的开关基因,仅有一个操纵基因。因此,有人开玩笑说:“半个操纵子就可以得诺贝尔奖”。对某一项成就,人们如果说它的一半就可以实现某种重要作用,就表明这项成就的伟大。北宋时代的名臣赵普就有“半部论语就可以治天下”的名言,由此也可见操纵子学说的巨大意义。
操纵子模型
一个控制细胞基因表达的模型称为操纵子(operon),此模型的提出使基因概念又向前迈出了一大步。表明人们已认识到基因的功能并不是固定不变的,而是可以根据环境的变化进行调节。随之人们发现无论是真核还是原核生物转录调节都是涉及到编码蛋白的基因和DNA上的元件。这一发现获得了1965年诺贝尔奖。一个基因就是一段编码有功能产物的DNA顺序。基因的产物可以是蛋白质或是RNA(如tRNA和rRNA)。基因的重要特点是在有的情况下其产物能从合成位点散开去作用别的位点。
DNA元件是DNA上一段顺序,它不能转变成任何其它的形式,但它作为一种原位(in situ)顺序具有特殊的功能。由于它只能作用同一条DNA,因此称顺式作用元件(cis-acting element)。
基因可以根据它们的产物分成不同的类型。编码细胞必要的蛋白,如酶或结构蛋白的基因称为结构基因( structural genes)。这类基因在细胞中占绝大部分,承担着细胞各种蛋白的结构和功能。编码调节蛋白的基因称调节基因(regulator genes)。调节蛋白可调节其它基因的表达。由于调节基因的产物可以自由地结合到其相应的靶上,因此被为反式作用因子(trans-acting因子)。
调节的关键是调节基因编码调节蛋白,此蛋白通过和DNA上特殊位点的结合来控制转录。顺式作用元件被反式作用因子识别可以以“正”的或“负”的形式调节靶基因,顺式作用位点通常总是在靶基因的上游。
操纵子学说相关的文章

菊目,拉丁学名Asterales,为显花植物的一目,仅菊科 一科,是显花植物 最大的科。分部广布全世界,热带较少,草本,半灌木或灌木,稀乔木,有乳汁管和树脂道,最显著特点是头状花序,多为一至多年生草本,生长于温带和亚热带的向阳处,分布范围从北极到南极,从海岸到树木线以上的高山。
科威特战争于1990年8月爆发,是伊拉克对科威特发动侵略战争,其意图将科威特变为伊拉克的第十九个省。五个月后,美国发动对伊拉克的战争,伊拉克不但被迫放弃了科威特,本国各项发展也遭到巨大破坏。

乌鲁木齐,维吾尔语:“ئۈرۈمچىشەھىرى”,简称乌市,旧称迪化,是中华人民共和国新疆维吾尔自治区辖地级市,首府城市,,位于新疆中北部。截至2022年末,乌鲁木齐市域面积13788平方千米,下辖7个市辖区、1个县,常住人口408.24万人。

尚可名片
这家伙太懒了,什么都没写!
作者


