
叶绿体基因组也叫叶绿体DNA(cpDNA),双链环状,每个叶绿体中约含12个cpDNA分子。叶绿体具有独立基因组,被认为是内共生起源的细胞器。
精选百科
本文由作者推荐
叶绿体基因组
环状双链DNA分子
外文名
The chloroplast genome
正文
(图)叶绿体基因组
叶绿体基因组是多拷贝的,具有比较保守的环状结构,但也存在着一些例外。叶绿体基因组主要用于编码与光合作用密切相关的一些蛋白和一些核糖体蛋白。叶绿体基因表达调控是在不同水平上进行的,光和细胞分裂素对叶绿体基因的表达也起着重要的调节作用。简介
(图)叶绿体基因组
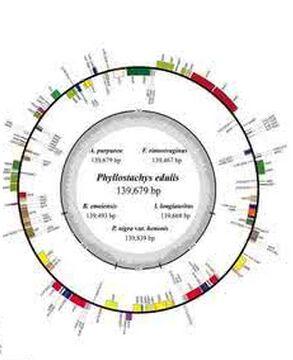
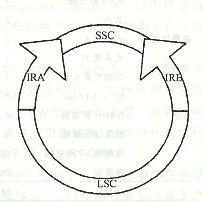
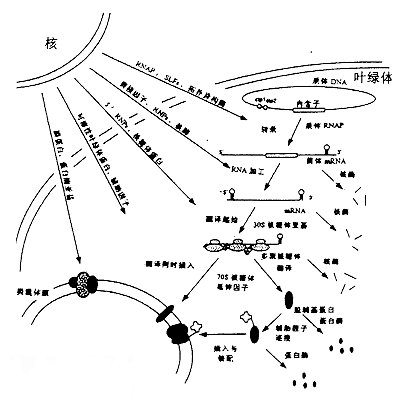
叶绿体是地球上绿色植物把光能转化为化学能的重要细胞器,叶绿体中进行的光合作用是严格地受到遗传控制的。早在20世纪初,人们就已知叶绿体的某些性状是呈非孟德尔式遗传的,但直到60年代才发现了叶绿体DNA(chloroplast DNA,cpDNA)。叶绿体基因组是一个裸露的环状双链DNA分子,其大小在120kb到217kb之间,相当于噬菌体基因组的大小,例如,T4噬菌体的基因组约165kb。一个叶绿体中通常有一个到几十个叶绿体基因组。叶绿体DNA不含5/—甲基胞嘧啶,这是鉴定ctDNA及其纯度的特定指标。叶绿体基因组中的基因数目多于线粒体基因组,编码蛋白质合成所需的各种tRNA和rRNA以及大约50多种蛋白质,其中包括RNA聚合酶、核糖体蛋白质、核酮糖1,5—二磷酸核酮糖羟化酶(RuBP酶)的大亚基等。高等植物的叶绿体基因组的长度各异,但均有10~24kb的一段DNA序列的两份拷贝,互呈反向重复序列(1RA和IRB)。这两份反向重复序列之间发生重组,形成了一份短的单拷贝序列(short single copy,SSC),把IRA和IRB连接起来,基因组的其余部分则是长的单拷贝序列(long single copy,LSC)。叶绿体基因组同线粒体基因组一样,都是细胞里相对独立的一个遗传系统。叶绿体基因组可以自主地进行复制,但同时需要细胞核遗传系统提供遗传信息。例如,光合系统Ⅱ中的chla/b蛋白质是在细胞质内的80S核糖体上合成后再转运进叶绿体的;RuBP酶的大亚基是在叶绿体内合成的,但其小亚基则是在细胞质中80s核糖体上合成后转运进叶绿体,然后同大亚基装配成有生物学活性的全酶。
cpDNA
(图)叶绿体基因组
叶绿体基因组在很多方面与线粒体基因组的结构是相似的。叶绿体DNA(cpDNA)是双链环状,缺乏组蛋白和超螺旋。cpDNA中的GC含量与核DNA及mtDNA有 很大的不同。因此可用CsCl密度梯度离心来分离cpDNA。每个叶绿体中cpDNA的拷贝数随着物种的不同而不同。但都是多拷贝的。这些拷贝位于类核区。例如甜菜的叶细胞中每个类核体有4~8个拷贝的cpDNA,而每个叶绿体有4~18个类核体,每个细胞中约有40叶绿体。每个细胞总共有约6000cpDNA分子。在衣藻中(chlamydomonas)(单细胞生物)在细胞中一个叶绿体含有500~1500 cpDNA分子。
烟草和水稻(Oryza sativa)叶绿体全序列分析表明cpDNA基因组成有以下特点:
1.基因组由两个反向重复序列(IR)和一个短单拷贝序列(short single copy seguence, SSC)及一个长单拷贝序列(long single copy seguence, LSC)组成;
2.IRA和IRB长各10-24Kb,编码相同,方向相反。
3.cpDNA启动子和原核生物的相似,有的基因产生单顺反子的mRNA,有的为多顺反子mRNA;
4.尽管cpDNA大小各不相同,但基因组成是相似的,而且所有基因的数目几乎是相同的,它们大部分产物是类囊体的成分或和氧化还原反应有关(表20-7);
5.其tRNA基因(IRA、IRB上各有7个,LSC上有23个,共37个)中有内合子存在,最长者达2526bp,此和原核tRNA不同。有的内合子位于D环上,此和原核tRNA不同。有的内含子位于D环上,此和真核生物核tRNA内含子常位于反密码子环上也不相同;
6.所有叶绿体基因转录的mRNA都由叶绿体核糖体翻译。
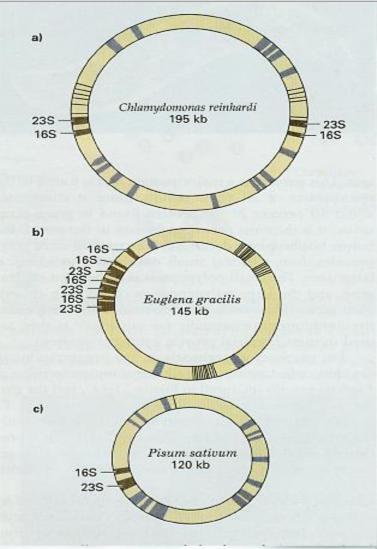
并不是所有的叶绿体都含有IR,IR上含有4种rRNA基因,根据它们排列的情况叶绿体可分为3类:I类是IR 序列,4种rRNA各有2个拷贝,对称分布在IR上cpDNA也较大,如玉米、烟草、水稻、菠菜、地钱、衣藻(C.Yreinhardi),大部分叶绿体都属此类。II类:无反向重复IR,而在cpDNA一侧16S,23S以正向串联重复的形式(各3个拷贝)排列。如少数低等植物,裸藻(Euglena gracilis);III类:无IR和DR,rRNA只有一拷贝,如豌豆(Posum satirum)等。这可能在进化的过程中DNA片段的重复和倒位而造成的。
相关研究
(图)叶绿体基因组
植物叶绿体基因组基因表达调控的研究
叶绿体基因组的特点是具相同或相关功能的基因组成复合操纵子结构。这一特点有利于叶绿体基因的表达与调控,例如rpoB-rpoC-rpoC 2操纵子是由编码RNA聚合酶各个亚基的基因聚合在一起而形成的,而psbI-psbK-psbD-psbC操纵子则编码PSⅡ的部分蛋白质。叶绿体基因组基因表达调控方式。
转录水平调节。转录后调节与修饰。莱茵衣藻核基因组与叶绿体基因组遗传转化体系的建立,以及许多光合途径缺陷突变体的分离为研究转录后调节提供了一个非常有用的模式系统。遗传分析表明RNA加工和RNA编辑为影响叶绿体基因表达转录后调节的因素。翻译水平调节。翻译水平调节可使生物快速地适应外界环境条件,特别对于高效表达基因,当环境条件不利时,可通过翻译水平快速调节,从而减少代谢能源的消耗。RNA水平和细胞器代谢状态影响叶绿体蛋白的翻译, 这种调节可能是通过核糖体蛋白反式磷酸化来完成的。翻译后调节与修饰。对于质体编码的叶绿素。
在每个叶原基细胞增殖过程中,位置信息决定细胞命运,因而不同细胞如叶肉细胞、皮层细胞、保卫细胞中对叶绿体的发育进行微调,大多数是通过调节RNA稳定性、剪接、翻译以及蛋白质稳定性来实现的,并显示核基因可以控制那些核和质体共同编码的、最终装配为复合体的蛋白基因。当发育为叶片时,不同细胞类型的核基因表达有所不同,不同细胞的位置信息,通过不同的基因调节机制,引起质体和核基因的细胞特异表达。最后,叶片细胞以关掉编码叶绿体蛋白的基因和核基因表达而进入衰老阶段。
基因表达调控是由一系列复杂的调控机制组成的。不同的调节机制在一定条件下对特定基因起调节作用,不同的调节策略可使不同植物来适应各自的生存条件,如:光、温、水和营养条件可调节植物的代谢活动。除上面提到的环境因素外,还涉及叶绿体基因转录及转录后调节、翻译与翻译后修饰调节、核基因对叶绿体基因在转录与翻译过程中的调节和质体产生的信号对核编码的质体蛋白的表达调节等等。因此,很难对叶绿体基因表达找出一个固定模式。在未来的研究中,核基因组和质体基因组如何在质体发育过程中起到相互调节作用将会成为一个最可能出成果的研究领域。
(图)叶绿体基因组
蓝藻和叶绿体基因组的比较研究
原核的蓝藻和真核植物(包括其他藻类)中的叶绿体,都同样进行放氧的光合作用,这为人类和整个生物界提供了赖以生存的食物、氧气、能源和原料。对叶绿体和蓝藻的细胞结构和分子生物学特性作分析,证明真核生物的叶绿体可能起源于蓝藻祖先的内共生。这使蓝藻在20多年来已成为光合作用研究的模式生物。
蓝藻基因组的作图和测序由日本Kazusa DNA研究所以S.Tabata博士领导的研究组,于1994年开始对集胞藻(Synechocystis sp. PCC6803)作分析,已于1996年完成。最近他们又基本完成了对鱼腥藻(Anabaena sp. PCC7120)的全序列测定。集胞藻6803的基因组大小为3,573,470bp,含有3168个编码蛋白的潜在基因,占全基因组87%。它的基因密度为1.1kb/基因,一个基因表达的产物平均长度为326个氨基酸残基,这些都是细菌基因组的典型数据。在3168个潜在基因中,1416个基因(45%)与已知的相似,尚有1752个基因(55%)需要鉴定。1416个已知基因中,按生物学功能可分成15类,其中与光合和呼吸有关的有131个,与转录有关的为24个,与翻译有关的144个。
把10种叶绿体的光合器蛋白和光合代谢中蛋白与蓝藻比较同一性发现,进化上差异越大,它们的同一性越差;在不同基因的同一性也有不同,如编码光合器的同一性较高,编码光合代谢的基因同一性差些。在编码光合器的蛋白中,光系统I和II反应中心的蛋白同一性较好。现在要做的是如何解释从蓝藻进化到叶绿体失去了绝大部分基因及为何在叶绿体进化中保留下来的蛋白在同一性上有这样的差异,从这些差异上能否得到启示来改造基因来提高光合作用效率。
参考资料
[1]生命经纬 http://www.biox.cn/content/20050705/24667.htm[2]百奥知 http://sou.bioknow.cn/html/pages/215/paper_30215.htm[3]生物通 http://www.ebiotrade.com/newsf/2001-4/L0141312048.htm
叶绿体基因组相关的文章

柠檬清新的香气,可以提神醒脑、振奋精神,缓解烦躁,净化空气。檬精精油 对于皮肤及身体也有很多积极的调理作用。柠檬精油中的柠檬烯特别有益于美白、收敛、平衡油脂分泌、治疗青春痘等油性皮肤症状。
本雅明·内塔尼亚胡(בִּנְיָמִין נְתַנְיָהוּ,Benjamin Netanyahu),男,1949年10月21日出生于以色列特拉维夫-雅法,毕业于麻省理工学院政治家、军人,第九、十三任以色列总理,兼任外交部长、国防部长、卫生部长、移民部长。
鸡肉炒饭是一种食物,制作原料主要有鸡肉、米、盐等。焯蔬菜是尤其是绿叶蔬菜时,要在水开后加少许盐和油,蔬菜焯好后马上倒入凉水里冰一下。这样会使绿色蔬菜颜色更好看,味道也好吃。

意识模糊一种以意识内容改变为主的意识障碍意识模糊:意识障碍的程度比嗜睡深,是一种以意识内容改变为主的意识障碍,表现为注意力减退,情感反应淡漠,定向力障碍,活动减少,语言缺乏连贯性,对周围环境的理解和判段低于正常水平,可有错觉,幻觉,躁动,精神错乱等,常见于急性重症感染的高热期。另有一种一兴奋性增高为主的意识模糊,伴有知觉障碍,称为谵妄,表现为定力丧失,感觉错乱,躁动。

尚可名片
这家伙太懒了,什么都没写!
作者
