
精选百科
本文由作者推荐
光合作用
绿色细胞中发生的生化反应
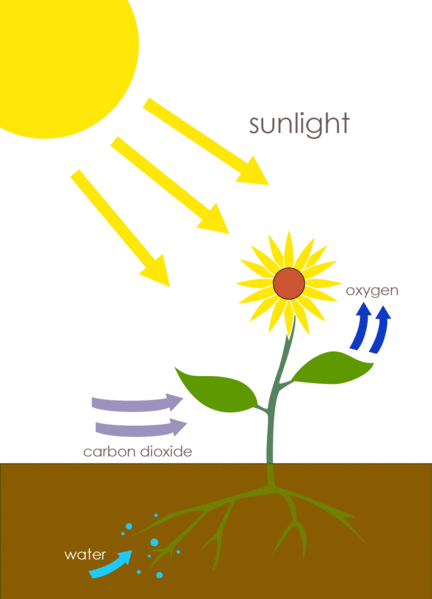
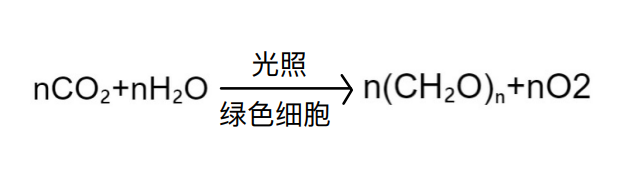
名称由来
地球上的生命都最终取决于来自太阳的能量,而光合作用又是唯一能收获这种能量的具有意义的过程。Photosynthesis(光合作用)一词字面意思便是利用光去合成(synthesis using light)。
演化过程
经过很长时间的演化,从这类微生物中衍生出了单细胞藻类那样的低等植物,可以进行释放氧气的光合作用。它们在水中进行光合作用,因此地表含量巨大的水无穷无尽地为生物提供着氧化底物,保证了地球上有机物质和氧气的大量积累,从而发展出了如今各种各样的生物。
光合作用的基因可能同源。罗伯特·布来肯细普团队利用BLAST检验了五种细菌的基因,发现其中有50个光合作用相关基因相似,因此他们对其进行亲缘关系分析,最终发现这些基因有着不同的演化史,只有少数同源基因,大多数的共同基因参与日常代谢反应,然后才被收纳称为光合系统的一部分。
发生场所
除植物外,部分光合细菌也可以进行光合作用。光合细菌(photosynthetic bacteria)是一类含有光合色素的原核生物,可以在厌氧光照条件下利用光能以二氧化碳或含碳有机物作为碳源进行不产生氧气的光合作用。
基本原理
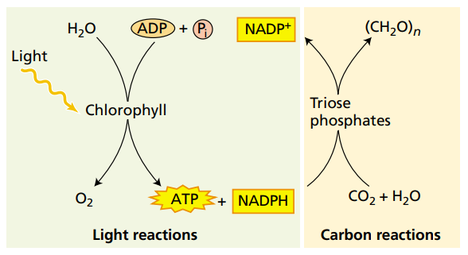
光合作用的光反应与暗反应(Taiz, Zeiger, 2010)
光反应
我们通常认为光反应由两个部分组成:原初反应阶段(吸收并传递光能,转化为电能)和同化力形成阶段(电子传递及其耦联的光合磷酸化完成)。
原初反应阶段
在此过程中,反应中心的叶绿素a分子作为原初电子供体受光子激发发生电荷分离(charge separation),失去电子被氧化,电子传递给原初电子受体,受体接收电子被还原,开始电子传递系列反应。光子的捕捉、传递过程由光合色素进行。
同化力形成阶段
同时,在与电子传递过程相耦联的光合磷酸化反应过程中,ADP和Pi在ATP合酶的催化下合成ATP。
在这两个过程中,形成的NADPH和ATP统称为同化力,用于下一阶段的反应进行。
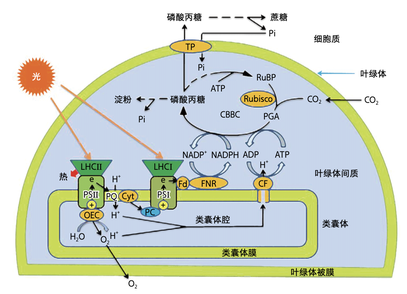
叶绿体中进行光合作用的过程模式图(许大全,2022)
暗反应
暗反应在叶绿体基质中发生,又称为卡尔文循环。在此过程中,ATP和NADPH在酶的催化下氧化成NADP和ADP+Pi,二氧化碳和水反应生成碳水化合物,也是我们所说的糖类物质。
光合碳循环分为以下三个阶段:
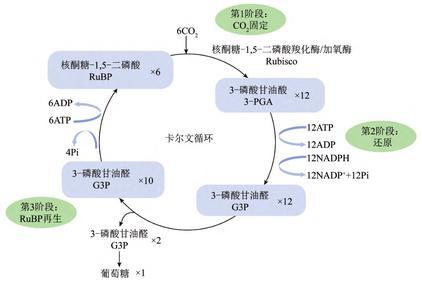
卡尔文循环示意图
羧化阶段:
6 RuBP + 6 CO2 

在CO2 浓度较低时,光合速率随着CO2 浓度的升高而直线升高,而这条直线的斜率也被称为羧化效率,同时羧化效率的大小可以判断RuBisCO量的多少。
在CO2 浓度较高时,光合速率不再随着CO2 的浓度升高而升高,此时的光合速率通常被称为叶片的光合能力,其大小依赖于RuBP再生能力的大小。
在CO2 浓度适中的过渡阶段,是从RuBP羧化限制到RuBP再生限制的转变阶段。
还原阶段
12 3-PGA + 12 ATP + 12 NADPH
还原阶段同时进行了磷酸化和NADPH2的还原。PGA是一种有机酸,若想转化为三碳糖(如磷酸甘油醛),需要使用NADPH2和ATP的同化力中的能量。当CO2被还原成三碳糖的水平时,光合作用的贮能部分便结束了。
再生阶段
2 G3P
10 G3P + 6 ATP
再生阶段是指通过一系列复杂的反应将RuBP再生出来,使CO2固定反应得以继续。再生的过程受到同化力供应的制约,因此需要光照。RuBP再生限制在某种程度上来说是可以反映光反应的情况,并了解光化学效率的。
注释
RuBP(ribulose bisphosphate)是一种在光合作用中依赖光的反应第一阶段形成的两个三碳中间体,是一种五碳糖。
不同生物的光合作用
C3途径植物
C3途径植物是一种具有特定碳同化途径的植物:二氧化碳最初固定产物是一种含有三个碳原子的有机物。C3途径就是我们熟悉的卡尔文循环,是美国加州大学的卡尔文(M. Calvin)及其同事于1946年通过C同位素标记法和双向纸层析技术得出并阐明的。C3途径发生在叶绿体基质中。
C4途径植物
二氧化碳最初固定产物是一种含有四个碳原子有机物的植物称为C4途径植物。C4途径又被称为Hatch and Slack Cycle,也被称为β-羧化途径和协同光合作用,是由澳大利亚植物生理学家、生化学家Marshall Davidson Hatch和英国植物生物学家、生化学家Charles Roger Slack研究甘蔗植物时得出的。在酶的催化下,二氧化碳和PEP(磷酸烯醇式丙酮酸)反应形成OAA(草酰乙酸),OAA在苹果酸脱氢酶的作用下转化为苹果酸。整个过程中,光反应发生在叶肉细胞中,而CO2同化作用发生在叶肉的维管束鞘细胞中。

C4植物种C4途径与C3途径的关系
CAM途径植物
CAM途径又称景天酸代谢途径,是一种最先在景天科植物中发现的CO2特殊固定方式,如今已在二十多科植物中发现该途径,如大戟科、百合科、仙人掌科、兰科、番杏科等,在裸子植物和蕨类植物中也有发现。常见的CAM途径植物有仙人掌、芦荟、龙舌兰、凤梨等。
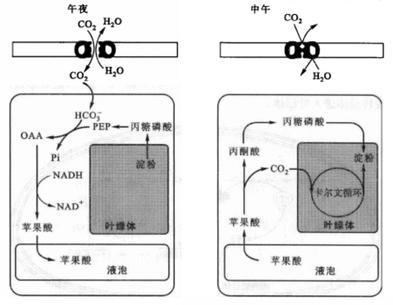
景天酸代谢示意图(潘瑞炽,2008)
特征
CAM植物在夜间开放叶片气孔来吸收二氧化碳,在夜间积累合成的苹果酸;而在白天,植物叶片气孔关闭防止水分散失,并将苹果酸转化为淀粉或其他化合物。
分类
CAM途径植物分为专一CAM和兼性CAM。专一CAM即使环境改变也会进行夜晚固定二氧化碳、积累有机酸来保证光合作用;而兼性CAM是在水分充足或幼苗时期表现为C3途径,在干旱炎热、水分匮乏或完全发育成熟时表现为CAM途径。兰科、番杏科、藤黄科、景天科、马齿苋科、葡萄科等中都存在兼性CAM植物,该途径帮助植物降低水分消耗,在不适宜的环境条件下保持了光合作用的完整性,得以生存繁殖。
藻类与细菌
在细菌和蓝绿藻类中,细胞质中的载色体上存在光合作用反应中心,组成了光合系统,其包含了细菌叶绿素分子以外,还包含了色素、细胞色素、含有非血红素铁和铜的分子、脂质、蛋白质等。这些生物体内有固氮酶或氢化酶对反应过程进行催化,从而合成氢气。
植物型光合作用
蓝藻进行植物型光合作用,采用环式电子传递的结构基础来提高卡尔文循环的效率,并能够帮助光和生物快速适应环境变化。
经固氮酶催化产氢反应式为: 2 H + Fd + 4 ATP = H2 + 4 ADP
细菌型光合作用
光合细菌暴露在光下时会诱导固氮酶激活,乳酸等有机物充当电子供体时,可以产生氢气。目前Rhodobacter sphaeroides已被认为是产氢率最高的细菌(可达260ml/mg/h),其光能转化效率为7%(意味着它产生的氢气燃烧所产生能量为吸收太阳能的7%),进一步开发可与太阳能电池的转化效率相当。
经固氮酶催化产氢反应式为:
C3H6O3 + 3 H2O = 12 H + 3 CO2 + 12 e = 6 H2 + 3 CO2
研究历史
在多国科学家的努力下,建立光合作用的化学方程式历时上百年。
参与反应气体的发现
最初在1771年,普里斯特利(Joseph Priestley)在研究气体时分离了八种气体,并描述了他们的特征。当时人们受到亚里士多德的教导,相信只有一种空气,而普里斯特利的实验证实说明了不同物质中释放不同空气。后针对实验结果展开可燃性原理的假说并付诸实验,将氧气称为了“去燃素空气(dephlogisticated air)”1777年,拉瓦锡(Antonie-Laurent Lavoiser)向科学院提交了《燃烧概论》的文章,将中一种气体称为“氧气(oxygen)”,解释了燃烧的本质。
光合作用的发现进程
1779年,出生于荷兰的英国医生科学家简·英格豪斯(Jan Ingehousz)发表了他关于绿色植物在阳光下可以吸收二氧化碳并释放氧气的发现,并指出绿色植物有在光照下净化普通空气的能力,这也是光合作用的雏形。在实验中,他将薄荷放入不含氧气的容器中,观察到几天后薄荷产生的物质使封闭的空气再次支持燃烧,这就是后来认为的氧气。并且阳光下,植物叶子浸泡在水中时,其绿色部分下面会出现小气泡,而在黑暗中不会,在非绿色部分也不会。同时,他还观察到在黑暗中植物释放二氧化碳气体“破坏空气”(后被认为是呼吸作用),但在光照下光合作用的速率远高于这种破坏作用的速率。
6 CO2 + 6 H2O 

20世纪中期,美国科学家卡尔文(Melvin Calvin)及其同事采用放射性同位素和色谱法等方式,通过追踪碳元素同位素,深入了解了光合作用中二氧化碳的应用与路径,并于1961年获得诺贝尔化学奖。这个过程现在被命名为“卡尔文循环”。
1954年,美国科学家Daniel Israel Arnon等人提出“同化力”,同时,发现叶绿体在光照下可以合成ATP,即发现了光合磷酸化的过程。
1957年,希尔(R. Hill)等人在爱默生(Emerson)关于光合效率的研究基础上提出了双光系统(two photosystem)的概念, 将小于或等于680nm波长的光系统定义为光系统II(PSII),将大于680nm波长的光系统称为光系统I(PSI)。同年,美国科学家D.I. Arnon又发现了光合磷酸化与Hill反应相偶联,再次论述了同化力的作用。
1966年,澳大利亚植物生理学家、生化学家Marshall Davidson Hatch和英国植物生物学家、生化学家Charles Roger Slack发现了一些植物叶肉细胞中产生四碳化合物,运送至维管束鞘细胞中以释放二氧化碳用于卡尔文循环,将这个途径命名为“Hatch and Slack pathway”。
1988年,德国科学家Johann Deisenhofer,德国生化学家Robert Huber和德国生化学家Hartmut Michel凭借对于光合反应中心的三维结构的确定获得诺贝尔化学奖。
1997年,美国生化学家Paul Boyer和英国生化学家John Walker因阐明ATP合成的酶促机制,共同获得1997年诺贝尔化学奖。
光子学说的提出
1905年,爱因斯坦提出了光子学说,并于1912年阐述了光化学当量定律。德国生理生化学家瓦博(Warburg)开始进行相应实验并得出与理论计算完全一样的结果。同时,瓦博的学生爱默生(R.Emerson)持反对意见,认为光合放氧的最低量子值为8。如今爱默生的观点获得广泛接受。
研究进展
发生于20世纪的一系列重大突破大大改变了光合作用研究的方向和范本,极大限度地综合了物理学、化学等学科理论与方法。
各国科学家对于光合作用相关主题的研究从未中断,如发现叶绿体蛋白降解新途径途径介导植物器官发育和抗逆境过程,仿生水裂解催化剂的研发,制作光合作用生物电池并应用于衰老动物细胞,与细胞色素c结合的酸杆菌同型二聚体反应中心的结构的发现等等。
相关名词
光合效率
光合效率(photosynthetic efficiency)指在一定时间范围内、一定光照强度下,光合作用反应的多少。每吸收一摩尔二氧化碳,则约有114千卡自由能储存在植物中。
光合色素
光呼吸
光呼吸(photorespiration),又称为乙醇酸氧化途径(glycollic acid oxidation pathway)或C2循环(C2 cycle),是植物的绿色细胞依赖光照吸收O2放出CO2的过程。该过程与光合作用密切相关,以C2化合物(乙醇酸)为底物,由叶绿体、过氧化物酶体、线粒体协同完成。一般生活细胞的呼吸对于光照没有特殊要求,即有无光照均可,称为暗呼吸。
意义影响
光合作用对于地球上绝大多数生命的存在至关重要。
制造有机物
如果光合作用停止,地球上很快就会没有食物或其他有机物。大多数生物便都会消失,地球的大气层很快就会变得几乎没有气态氧。因此,人们把地球上的绿色植物比作“绿色工厂”,绿色植物的生存离不开自身进行光合作用产生的有机物,而人类和动物的食物也直接或间接地来自光合作用产生的有机物。
转化并储存太阳能
数百万年前,植物进行光合作用产生的能量还负责为工业社会提供化学燃料,即煤、石油、天然气,地球上几乎所有生物都直接或间接地利用着这些能量。这些燃料中含有的能量都是被古代植物通过光合作用储存起来的,只是现代文明将这些曾经从空气中去除的CO2又返还回去,预计这一现象将对地球气候产生重大影响。
使大气中碳氧平衡
在人口快速增长的现代,对食物、材料、能源需求的增长,导致了对光合作用产物转化为对人类有用的产品的效率需求增长。而如今园林、城规建设中的碳氧平衡法也旨在解决当前问题,以平衡城市中居民呼出及其他燃质产生的CO2,以达到大气中碳氧平衡。
对生物进化有重要作用
光合作用产生氧气和糖类,为有氧呼吸作用的生物创造了便利条件,提供能量供生物使用。并且氧气在太阳光紫外线的照射下会形成高空臭氧层(O3),有效防止紫外线直接照射地球表面的生物造成伤害,保证生物圈的正常形成与运作。
光合作用相关的文章

阿尔泰山脉(英语:Altai Mountains;俄语:Алтай),是位于欧亚大陆中部的山系,坐落在北纬45°~52°,东经84°~99°之间,地跨哈萨克斯坦、俄罗斯、中国和蒙古四国,整体呈西北-东南走向。其东北与萨彦岭相连,随后地势向东南逐渐变低,并与戈壁沙漠相接。该山脉东西总长约2000千米,
亲历中国科幻内容介绍本丛书的编辑宗旨不是面面俱到,而是像丛书名字中所陈述的,期望在新思维方面给科幻研究领域一些启迪。阅读中读者一定会发现,无论从各分册或文章的选题还是从研究方法上,本丛书的作者都突破了以往比较单一的文学研究模式,试图以多元视角和方法探索科幻文学。笔者认为,对于科幻这种跨越学科的文学存

尚可名片
这家伙太懒了,什么都没写!
作者


