精选百科
本文由作者推荐
蛋白激酶
催化蛋白质磷酸化反应的酶
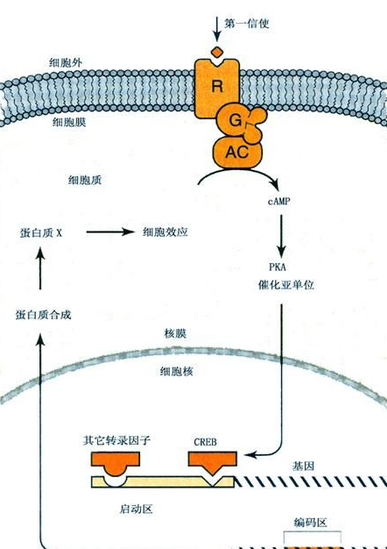
蛋白激酶(protein kinases,简称PK)。催化蛋白质磷酸化过程的酶。蛋白质的磷酸化过程是神经信息在细胞内传递的最后环节.导致离子通道蛋白及通道门的状态变化。在神经细胞内有许多种类。在半静状态下,处于小活动状态。根据其活化条件的不同,可分为蛋白激酶A、蛋白激酶G、蛋白激酶C和钙调蛋白的蛋白激酶。蛋白激酶A,也称环腺苷酸依存性蛋白激酶.只有第二信使环腺苷酸才能激活它。主要存在于大脑皮层和纹状体。蛋白激酶B,只有有了环磷酸腺苷,才能激活,并接替环磷酸腺苷在细胞内传递信息。在脑内含量少,且难以与蛋白激酶A分离。其在脑内的特异性分布,至今尚不清楚。蛋白激酶C可被二酰甘油(DG)激活。钙调蛋白的蛋白激酶可被钙离子和钙调蛋白激活。
研究历史
种类
到目前为止,已发现的蛋白激酶约有400多种,分子内都存在一个同源的由约270氨基酸残基构成的催化结构区。在细胞信号传导、细胞周期调控等系统中,蛋白激酶形成了纵横交错的网络。这类酶催化从ATP转移出磷酸并共价结合到特定蛋白质分子中某些丝氨酸、苏氨酸或酪氨酸残基的羟基上,从而改变蛋白质、酶的构象和活性。
蛋白质磷酸化反应是指三磷酸腺苷(ATP)末位(γ位)的磷酸转移到基质蛋白质的特定氨基上所进行的共价修饰的一类反应的总和。蛋白激酶催化这类反应。可分为环腺苷酸(cAMP)-依赖性蛋白激酶:环鸟苷酸(cGMP)-依赖性蛋白激酶、钙调素依赖性蛋白激酶和对磷脂敏感的钙离子依赖性蛋白激酶。一般由蛋白激酶催化磷酸化反应中接受磷酸基的部位是丝氨酸、苏氨酸或酪氨酸的羟基。
蛋白激酶很多,根据其底物蛋白被磷酸化的氨基酸残基种类,可将它们分为5类,即:
①丝氨酸/苏氨酸(Ser/Thr)蛋白激酶:蛋白质的羟基被磷酸化;
②酪氨酸(Tyr)蛋白激酶:蛋白质的酚羟基作为磷受体;
④色氨酸蛋白激酶:以蛋白质的色氨酸残基作为磷受体;
⑤天冬氨酰基/谷氨酰基蛋白激酶:以蛋白质的酰基为磷受体。
目前发现的植物蛋白激酶以前3类为主。而Stone和Walker(1995)根据蛋白激酶催化区域氨基酸序列的相似性,将植物蛋白激酶分为5大组。这5大组蛋白激酶分别为①AGC组:以cAMP(环腺苷酸)依赖的蛋白激酶PKA、cGMP(环鸟苷酸)依赖的蛋白酶PKG及钙和磷脂依赖的蛋白激酶PKC为代表,以受第二信使(如cAMP、cGMP、DAG(二酰甘油)和Ca2+)激活为特征。②CaMK组:包括Ca2+/CaM依赖的蛋白激酶CaMK、Ca+依赖而CaM不依赖的蛋白激酶CDPK等,依赖第二信使是该组蛋白激酶的普遍性。③CMGC组:包括MAPK(分裂原激活的蛋白激酶)、CDK(周期素依赖的蛋白激酶)等,相对于前2组蛋白激酶依赖于第二信使,该组激酶作用于下游的磷酸化级联系统。④传统的PTK组:为酪氨酸蛋白激酶,目前在植物中尚未发现纯粹的酪氨酸蛋白激酶,但并不意味着Tyr残基的磷酸化对植物不重要。二重特异性蛋白激酶如MAPKK在植物中的发现,证明了Tyr残基的磷酸化可能在高等植物中具有重要的生理作用。⑤其它组:如类受体蛋白激酶RLKs及乙烯信号转导元件CTRl(胞质级联蛋白激酶MAPKKK)等。
蛋白激酶A(Protein kinase A,简称PKA),也称为环磷酸腺苷依赖蛋白激酶(cAMP-dependent protein kinase、简称cAPK)。是一种酶,其活性依赖于细胞中环磷酸腺苷(cAMP)的含量。
PKA是一个全酶(holoenzyme,由许多次单位组成,是完整的且有作用的酶),它包含了两个调控次单位以及两个代谢次单位。当细胞中的cAMP较少时,PKA虽然会暂时失去活性,但仍然可以保持结构完整。当cAMP浓度增加,cAMP会接上位于两个调控次单位上的活性区(Binding site),并使蛋白激酶A的构形改变,进而将两个代谢次单位释放。自由的代谢次单位,则可以参与一些化学反应。
分布
蛋白激酶在细胞内的分布遍及核、线粒体、微粒体和胞液。一般分为3大类。①底物专一的蛋白激酶:如磷酸化酶激酶,丙酮酸脱氢酶激酶等。②依赖于环核苷酸的蛋白激酶:如环腺苷酸(cAMP)蛋白激酶,环鸟苷酸(cGMP)蛋白激酶。③其他蛋白激酶:如组蛋白激酶等。
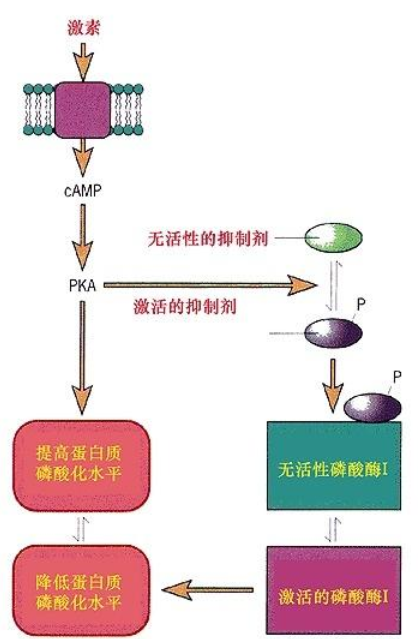
cAMP蛋白激酶以活化型和非活化型两种形式存在于生物体中,它们的比例受到多种激素的调节控制。cAMP蛋白激酶是由4个亚基组成——两个能与cAMP结合的调节亚基(R)和两个非活化的能催化磷酸基团转移的催化亚基(C)。如果细胞内cAMP浓度升高则cAMP蛋白激酶解离成调节双亚基和两个活化的催化亚基(见图)。
蛋白激酶相关的文章

三氯甲烷,分子式为CHCl3,为无色透明液体,有特殊气味,味甜,高折光,不燃,质重,易挥发。对光敏感,遇光照会与空气中的氧作用,逐渐分解而生成剧毒的光气(碳酰氯)和氯化氢。可加入0.6%~1%的乙醇作稳定剂。能与乙醇、苯、乙醚、石油醚、四氯化碳、二硫化碳和油类等混溶、25℃时1mL溶于200mL水。
灵芝又称林中灵、琼珍(学名:Ganoderma Lucidum Karst)是多孔菌科真菌灵芝的子实体。具有补气安神、止咳平喘、延年益寿的功效。用于眩晕不眠、心悸气短、神经衰弱、虚劳咳喘。其外形呈伞状,菌盖肾形、半圆形或近圆形。在明代大医学家李时珍编著的《本草纲目》中有灵芝的记载。
柠檬精,网络流行词。其字面意思就是柠檬成精,用来表示自己很酸的感觉,指很喜欢酸别人,嫉妒别人,指嫉妒心很强,见不得别人比自己好,又自视清高的那种人。后来其贬义色彩逐渐淡化,多用于自嘲式地表达对他人从外貌到内在、从物质生活到情感生活的多重羡慕。
西班牙王国(西班牙语:Reino de España;英语:The Kingdom of Spain),简称“西班牙”,是主要位于欧洲西南部的君主立宪制国家。 截至2023年,西班牙总人口为4761.5万,国土面积约为50.6万平方千米,海岸线长约7800千米,领海面积为43332平方千米。西班牙的

尚可名片
这家伙太懒了,什么都没写!
作者