
精选百科
本文由作者推荐
乙酰胆碱
调控生殖机能的神经递质
基本信息
| 中文名 | 乙酰胆碱 |
| 外文名 | ACh, acetylcholine |
| 别名 | ACh AcCh |
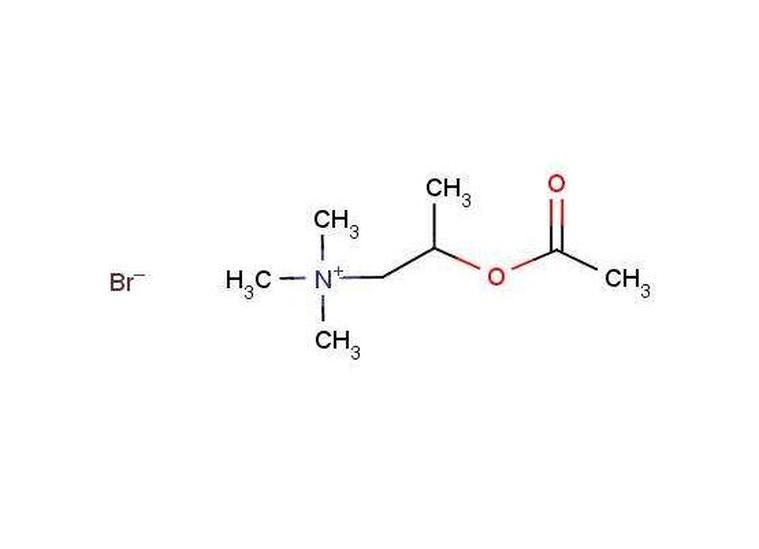
| 化学式 | CH3COOCH2CH2N+(CH3)3 |
| 外观 | 神经递质之一 |
展开
简介
乙酰胆碱的认识
1914年,Ewins在麦角菌中发现了乙酰胆碱,这是首次在非神经细胞中发现乙酰胆碱的报道。随后,人们陆续在多种细菌、真菌、低等植物和高等植物中发现了乙酰胆碱及其相关的酶和受体。随着胆碱能系统在植物中的发现和研究的深入,人们似乎有望在分子水平发现动植物间的又一相似性,因而植物学家抱着极大的热情投入了这方面的研究。但是由于当时研究手段的限制、对动植物之间的差别认识不足,以及某些研究在其它的实验室难以重复的缘故,使得植物乙酰胆碱的研究多处于零星的、非系统的状态,研究的深度和广度远远无法与动物相比。尚未对其在植物中的作用机理提出一个合理的解释。近年来,我们和国外其它几家实验室重新开展了乙酰胆碱在植物体内的生理作用和作用机理的研究,为揭示植物乙酰胆碱的作用机理提供了新的线索。
作用
植物生理过程
种子萌发
乙酰胆碱和乙酰胆碱酯酶可能参与调控某些植物的种子萌发和幼苗早期生长,乙酰胆碱影响这些生理过程的机理可能涉及调控储藏物从下胚轴向植物快速生长部位的调运。乙酰胆碱对需光种子萌发的影响的研究有许多矛盾的报道。Tretyn等在研究乙酰胆碱及其类似物、乙酰胆碱酯酶抑制剂对不同光周期植物种子萌发的影响中发现,无论在光下还是在黑暗中这些化合物对光不敏感植物的种子萌发都没有影响。但在光下可以促进需光种子萌发,而在暗中则没有作用。对于不需光种子,乙酰胆碱抑制其在光下的萌发,乙酰胆碱类似物胆碱对上述过程则无影响。由于乙酰胆碱及乙酰胆碱酯酶在种子中广泛存在,因而有理由推测乙酰胆碱参与调控种子的萌发,调控的机理可能涉及光对种子中乙酰胆碱酯酶活性的抑制。
生长
乙酰胆碱对于生长的影响因实验条件的不同,植物种类或同一植物不同组织而异。乙酰胆碱可以模拟红光的作用抑制大豆侧根的发育,还可以引起小麦幼苗生长和干重的增加。在离体组织中,乙酰胆碱可以刺激燕麦胚芽鞘和黄瓜下胚轴的伸长以及绿豆下胚轴的生长,刺激蚕豆下胚轴的生长而抑制其上胚轴的生长。总之,乙酰胆碱对植物生理过程的影响与所利用的组织及实验条件密切相关,其最大效应在pH酸性区。
成花作用
乙酰胆碱可以模拟红光的作用,抑制远红光刺激的过氧化物酶活性升高,从而使菠菜能在非诱导条件下开花。乙酰胆碱可以抑制连续光照条件下(24 h光照/0 h黑暗)长日浮萍G1的成花和刺激非诱导的短日条件下短日浮萍Torr的成花。阿托品可以抑制连续光照下生长的青萍G3成花而管箭毒则无作用,说明乙酰胆碱对成花的诱导作用可能是通过质膜上的类毒蕈碱型受体介导的。乙酰胆碱对成花的诱导作用还可能与它调控的膜对离子的通透性有关。光周期诱导的成花作用涉及到叶片膜电势的改变,乙酰胆碱还可能通过影响膜电势而参与成花诱导。
呼吸作用
乙酰胆碱可引起根尖细胞耗氧速率的增加。Jaffe以游离的线粒体为材料得到的结果已证实了这一点。伴随着氧的消耗,组织中ATP的水平下降10倍,自由磷水平升高14倍。乙酰胆碱的这种作用可能是其使呼吸的电子传递链与氧化磷酸化解偶联所造成。根据这些实验结果,Jaffe提出了乙酰胆碱对大豆根尖细胞的作用模式,即当胞间乙酰胆碱浓度升高时,乙酰胆碱到达其作用的靶部位,随后是分泌质子,氧的消耗和ATP水解增加,而这些过程均与膜对阳离子通透性的增加相关连。
光合作用
棚田效应
红光促使黄化的绿豆和大麦根尖吸附到带负电的玻璃杯内壁上,而远红光则使根尖脱离杯壁释放到溶液中。这种现象称为棚田效应(tanada effect)。在黑暗中乙酰胆碱可以使离体的大豆根尖吸附到带负电的玻璃杯内壁上,并阻止远红光引起的根尖脱离杯壁,乙酰胆碱酯酶抑制剂毒扁豆碱(eserine)增加组织对乙酰胆碱的敏感性。这些说明内源乙酰胆碱可能在这一生理过程中有控制作用。
通透性
乙酰胆碱可以刺激质子从大豆根尖细胞流出,诱导菠菜叶片膜电势的变化,抑制蓝光诱导的大豆下胚轴弯钩膜电势的超极化及该组织对钾的吸收,这些过程都涉及乙酰胆碱对膜透性的调节。
除了影响上述过程外,乙酰胆碱还可以影响组织对钙离子的吸收。Tretyn发现乙酰胆碱可以刺激黄化燕麦胚芽鞘对钙离子的吸收。乙酰胆碱酯酶的抑制剂可以增加组织对乙酰胆碱的敏感性;钙通道的抑制剂可以抑制乙酰胆碱刺激的钙吸收的增加。这些结果表明乙酰胆碱参与调控植物的钙通道。
参与相互作用
参与植物与植物以及细胞与细胞之间的相互作用
在一个生态环境中,植物与植物之间以及植物与其他生物之间常常表现出相互作用的关系。这种相互作用可以是促进性的也可以是抑制性的,即表现为相生相克的关系。乙酰胆碱酯酶存在于根瘤菌感染大豆所形成的根瘤中,而且乙酰胆碱酯酶的最大活性与根瘤对氮的最大同化期相一致,推测乙酰胆碱及其酯酶在根瘤菌和寄主植物间的相互作用中起一定作用。乙酰胆碱酯酶还存在于地衣的叶状体中,而且主要分布于组成地衣的真菌和藻类两种生物的界面。在其粉芽(soredia)产生孢子过程中,乙酰胆碱酯酶活性增加,而且酶活性集中分布在接触区。乙酰胆碱及乙酰胆碱酯酶参与地衣这种生物间相互作用的机理可能是通过调控膜对离子的通透性,并介导环境中光对地衣生殖影响而实现的 乙酰胆碱及乙酰胆碱酯酶还可能参与花粉与柱头间的识别作用。在裂叶牵牛中乙酰胆碱酯酶主要分布在雌蕊柱头的表面,还存在于花粉粒和花粉管的尖端。乙酰胆碱的激活剂和拮抗剂以及乙酰胆碱酯酶的抑制剂均可以影响某些植物的花粉萌发和花粉管伸长。因此推测乙酰胆碱和乙酰胆碱酯酶可能参与植物柱头和花粉间的相互作用。
信号转导
原生质体膨胀 红光可以刺激黄化小麦叶肉细胞原生质体体积膨胀,这种刺激作用可为随后的远红光照射所逆转,说明这一反应是在光敏素控制下进行的。红光对原生质体体积膨胀的刺激作用要求介质中含有Ca。乙酰胆碱可以代替红光在黑暗中引起原生质体的膨胀。与红光引起的反应不同,乙酰胆碱不仅可以在含Ca的介质中引起原生质体的膨胀,而且在含Na或K的介质中也可以引起原生质体的膨胀。
除乙酰胆碱外,只有氨基甲酰胆碱可以刺激原生质体的膨胀,而胆碱、丙酰胆碱和丁酰胆碱则无此作用。乙酰胆碱酯酶的抑制剂毒扁豆碱可以增加原生质体对乙酰胆碱的敏感性。据此可以认为乙酰胆碱能特异地刺激黄化小麦叶肉原生质体膨胀
乙酰胆碱诱导原生质体膨胀过程中是否涉及乙酰胆碱受体的参与可用乙酰胆碱受体的激活剂和抑制剂来确定。乙酰胆碱N型受体的激活剂烟碱在含Na或K的介质中可以直接刺激原生质体膨胀,而在含Ca的介质中,烟碱没有作用。与以上结果不同,M型受体的激活剂毒蕈碱可以在含Ca的介质中刺激原生质体膨胀,而在含Na或K的介质中没有作用。采用乙酰胆碱受体抑制剂的研究也得出同样的结论。M型受体的抑制剂阿托品在含Na或K的介质中对乙酰胆碱刺激的原生质体膨胀没有作用,但在含Ca介质中则可以抑制乙酰胆碱诱导的原生质体膨胀。N型受体抑制剂管箭毒在含Ca介质中对乙酰胆碱刺激的原生质体膨胀没有作用,但在含Na或K的介质中则有抑制作用。荧光定位技术证明N型乙酰胆碱受体主要分布在原生质体表面。
在乙酰胆碱诱导的原生质体膨胀过程中,乙酰胆碱为受体接受后的信号转导可能涉及到Ca和CaM,因为Ca通道抑制剂硝苯地平(nifedipine, NIF)和La可以完全抑制乙酰胆碱诱导的原生质体在含Ca介质中的膨胀。同样,钙调素的抑制剂和G蛋白的抑制剂也有这样的作用,而这些化合物在含Na或K的介质中则没有作用。
幼叶展开 生长于黑暗中8 d的小麦幼苗,其初生叶的展开受控于光敏色素系统。如果介质中含有Ca,乙酰胆碱在暗中可以刺激离体叶切段中幼叶的展开。在没有Ca而有Na的介质中乙酰胆碱也可以刺激黄化小麦初生叶片的展开。在乙酰胆碱的各种衍生物中只有氨基甲酰乙酰胆碱可以刺激黄化小麦初生叶片的展开。乙酰胆碱受体的拮抗剂,阿托品和D-管箭毒可以分别抵消乙酰胆碱在含Ca和Na介质中诱导叶片的展开。乙酰胆碱受体的激活剂,毒蕈碱和烟碱可以分别在Ca和Na的介质中刺激原生质体的膨胀。乙酰胆碱诱导的Ca依赖的叶片开展可为Ca通道抑制剂硝苯地平和钙调素抑制剂3-氟-甲基吩噻嗪(trifluoperazine, TFP)所减弱,其中只有钙调素抑制剂TFP可以抑制乙酰胆碱诱导的在含Na介质中黄化小麦初生叶片的展开。
酶活性
乙酰胆碱在植物中的作用机理除参与调节膜对离子的通透性外,可能还涉及对植物体内某些酶活性的调控。乙酰胆碱对兵豆(Lens culinaris)根生长的抑制作用与体内过氧化物同工酶的活性变化密切相关,它可以刺激某些同工酶的活性而抑制另外一些同工酶的活性。
乙酰胆碱本身对于植物体内苯丙氨酸氨基裂解酶的活性和类黄酮的合成没有影响,但它却可以抵消红光对苯丙氨酸氨基裂解酶活性和类黄酮合成的刺激作用。
生理活动影响
中枢胆碱能系统与学习、记忆密切相关,乙酰胆碱(ACh)是中枢胆碱能系统中重要的神经递质之一,其主要功能是维持意识的清醒,在学习记忆中起重要作用。
人体利用
人的脑组织有大量乙酰胆碱,但乙酰胆碱的含量会随着年龄的增加而下降。正常老人比青年时下降30%,而老年痴呆患者下降更为严重,可达70%~80%。美国医生伍特曼观察到老年人脑组织乙酰胆碱减少,就给老年人吃富含胆碱的食品,发现有明显的防止记忆减退的作用。英国和加拿大等国的科学家也相继进行了研究,一致认为只要有控制地供给足够的胆碱,可避免60岁左右老年人记忆力减退。所以保持和提高大脑中乙酰胆碱的含量,是解决记忆力下降的根本途径。在自然界是,乙酰胆碱多以胆碱的状态存在于蛋、鱼、肉、大豆等之中,这些胆碱必须在人体内起生化反应后,才能合成具有生理活性的乙酰胆碱。另外,经常服用蜂王浆可以提高脑内乙酰胆碱的含量,从而促进激活脑神经传导功能,提高信息传递速度,增强大脑记忆能力,全面改善脑功能,并能延缓衰老。
药理作用
心血管系统
(1)血管扩张作用:静注小剂量本品可由于全身血管扩张而造成血压短暂下降,并伴有反射性心率加快。ACh可引起许多血管扩张。如肺和冠状血管。其扩血管作用主要由于激动血管内皮细胞M,胆碱受体亚型,导致内皮依赖性舒张因子(EDRF)即一氧化氮(nitric oxide,No)释放,从而引起邻近平滑肌细胞松弛,也可能通过压力感受器或化学感受器反射引起。如果血管内皮受损,则ACh的上述作用将不复存在,相反可引起血管收缩。此外,ACh通过激动交感神经末梢突触前膜M1受体,抑制去甲肾上腺素能神经末梢释放NA也与ACh扩血管作用有关。
(2)减慢心率:亦称负性频率作用。ACh能使窦房结舒张期自动除极延缓,复极化电流增加,使动作电位到达阈值的时间延长,导致心率减慢。
(3)减慢房室结和普肯耶纤维传导:即为负性传导作用。ACh可延长房室结和普肯耶纤维(Βurkinje fibers)的不应期,使其传导减慢。当使用强心苷使迷走神经张力增高或全身给药法使用大剂量胆碱受体激动药时所出现的完全性心脏传导阻滞常与房室结传导明显抑制有关。
(4)减弱心肌收缩力:即为负性肌力作用。一般认为胆碱能神经主要分布于窦房结、房室结、普肯耶纤维和心房,而心室较少有胆碱能神经支配,故ACh对心房收缩的抑制作用大于心室。但由于迷走神经末梢与交感神经末梢紧密相邻,迷走神经末梢所释放的ACh可激动交感神经末梢突触前M胆碱受体,反馈性抑制交感神经末梢去甲肾上腺素释放。使心室收缩力减弱。
泌尿道
相关合集

其他的神经递质
共5个词条1241阅读

1,4-丁二醇
可燃的无色粘稠油状液体
乙酰胆碱
调控生殖机能的神经递质
1,4-丁内酯
无色油状液体
查看更多
相关视频
全部
1699次播放02:14
乙酰胆碱是什么
乙酰胆碱相关的文章

脑炎是指脑实质受病原体侵袭导致的炎症性病变。绝大数的病因是病毒,也可由细菌、霉菌、螺旋体、立克次氏体、寄生虫等感染引起。脑炎的发病多因机体抵抗力下降,病原菌趁机通过呼吸道,消化道,性传播和密切接触等各种形式,经过血循环,透过血-脑屏障进入到大脑,或是脑组织邻近部位感染直接侵犯脑组织,从而引起脑炎的发
江苏省,简称“苏”,是中华人民共和国省级行政区,省会南京,位于长江三角洲地区,中国大陆东部沿海,地跨北纬30°45'~35°08',东经116°21'~121°56',与上海市、浙江省、安徽省、山东省接壤。江苏总面积10.72万平方公里。国家统计局数据显示,2021年江苏省常住人口为8505.4万人
小编整理:平冈佑太是一位日本男演员,出生于1984年9月1日,来自日本山口县。他在2002年参加JunonSuperBoyContest并获得冠军,从而开始进入娱乐界。以后来,他出演了多部电视剧,包括《借着雨点说爱你》和《东京friends》等。而在电影方面,
穆罕默德·穆尔西(Mohamed Morsi,1951年8月20日-2019年6月17日),全名穆罕默德·穆尔西·伊萨·阿耶特,政治家,埃及第5任总统,首位民选总统,毕业于美国南加利福尼亚大学工程学博士学位。


尚可名片
这家伙太懒了,什么都没写!
作者
