
精选百科
本文由作者推荐
聚合酶
1957年美国发现的物质
基本信息
| 中文名 | 聚合酶 |
| 外文名 | polymerase |
| 属性 | 蛋白质 |
| 发现时间 | 1957年 |
| 又称 | DNA聚合酶 |
| 概述 | 催化合成DNA核糖核酸一类酶 |
| 发现者 | 阿瑟·科恩伯格 |
收起
介绍
分类
聚合酶医学用途
特性
聚合作用
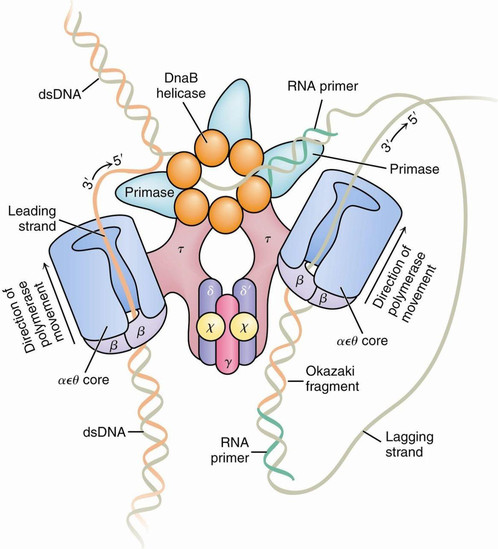
聚合酶结构图
3'5'外切酶活性──校对作用
这种酶活性的主要功能是从3'→5'方向识别和切除不配对的DNA生长链末端的核苷酸。当反应体系中没有反应底物dNTP时,由于没有聚合作用而出现暂时的游离现象,从而被3'→5'外切酶活性所降解。如果提高反应体系的温度可以促进这种作用,这表明温度升高使DNA生长链3'末端与模板发生分离的机会更多,因而降解作用加强。当向反应体系加入dNTP,而且只加放与模板互补的上述核苷酸才会使这种外切酶活性受到抑制,并继续进行DNA的合成。由此推论,3'→5'外切酶活性的主要功能是校对作用,当加入的核苷酸与模板不互补而游离时则被3'→5'外切酶切除,以便重新在这个位置上聚合对应的核苷酸。在某些T4噬菌体突变株中DNA复制的真实性降低,而易发生突变,从此突变株分离得到的聚合酶的3'→5'外切酶活性很低。相反,另外一些具有抗突变能力的T4突变株中的聚合酶的3'→5'外切酶活性比野生型高得多,因此,其DNA复制真实性好,变异率低。可见,3'→5'外切酶活性对DNA复制真实性的维持是十分重要的。
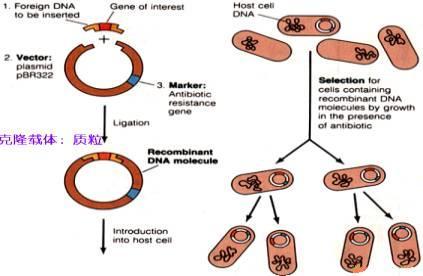
聚合酶分子反应示意图
5'→3'外切酶活性就是从5'→3'方向水解DNA生长链前方的DNA链,主要产生5'-脱氧核苷酸。这种酶活性只对DNA上配对部份(双链)磷酸二酯键有切割活力作用,方向是5'→3'。每次能切除10个核苷酸,而且DNA的聚合作用能刺激5'→3'外切酶活力达10倍以上。因此,这种酶活性在DNA损伤的修复中可能起着重要作用。对冈崎片段5'末端DNA引物的去除依赖此种外切酶活性。
焦磷酸解作用
DNApolⅠ的这种活性可以催化3'末端焦磷酸解DNA分子。这种作用就是无机焦磷酸分解DNA生长链,可以认为是DNA聚合作用的逆反应,而且这种水解DNA链作用需要有模板DNA的存在。(dNMP)n XPPi←(dNMP)n-x X(dNPPP)→DNA
焦磷酸交换作用
催化dNTP末端的PPi同无机焦磷酸的交换反应。反应式为32P32Pi dNPPP←dNP32P32P PPi→DNA
最后两种作用,都要求有较高浓度的PPi,因此,在体内由于没有足够高的PPi而无重要意义。DNApolⅠ的DNA聚合酶活性和5'→3'外切酶活性协同作用,可以使DNA链上的切口向前推进,即没有新的DNA合成,只有核苷酸的交换。这种反应叫缺口平移(Nicktranslation)。当双链DNA上某个磷酸二酯键断裂产生切口时,DNApoIⅠ能从切口开始合成新的NDA链,同时切除原来的旧链。这样,从切口开始合成了一条与被取代的旧链完全相同的新链。如果新掺入的脱氧核苷酸三磷酸为α-32P-dNTP,则重新合成的新链即为带有同位素标记的DNA分子,可以用作探针进行分子杂交实验。
尽管DNApolⅠ是第一个被鉴定的DNA聚合酶,但它不是在肠杆菌中DNA复制的主要聚合酶。主要证据如下:纯化的DNApolⅠ催化dNTP掺入的速率为667碱基/分,而体内DNA合成速率要比此高二倍数量级;大肠杆菌的一个突变株中,此酶的活力正常,但染色体DNA复制不正常;而在另一些突变株中,DNApolⅠ的活力中只是野生型的1%,但是DNA复制却正常,而且此突变株增加了对紫外线、烷化剂等突变因素的敏感性。这表明该酶与DNA复制关系不大,而在DNA修复中起着重要的作用。
一些特定的DNA聚合酶对于化学修饰性核苷分子显示出惊人的耐受性,从而为高度功能化的核酸分子的有效合成提供了令人激动的新机遇。
研究成果

聚合酶结构示意图
聚合酶分子反应示意图
现状
显微镜下的聚合酶
迄今为止,在真核细胞中已至少发现了12种DNA聚合酶,包括聚合酶α、β、γ、δ、ε、ζ、η、θ、ι、κ、λ和μ,它们通过参与DNA聚合反应核酸外切酶的校读作用或DNA修复过程,对DNA复制保真度的维持起重要作用,直接影响着细胞遗传的稳定性。
聚合酶相关的文章

世界气象组织(英语全称:World Meteorological Organization,简称:WMO),是联合国的一个专门机构,总部设于瑞士日内瓦,有193个会员国和会员地区(截至2023年6月) ,致力于在地球大气状态和变化规律及其与陆地和海洋的相互作用、大气产生的天气和气候、以及由此产生的水
亚洲(英语:Asia),是世界七大洲之一,以及亚欧大陆的主体部分,其西邻欧洲和非洲,东和南面太平洋和印度洋,最北抵北冰洋,总面积约4400万平方千米,囊括48个国家,总人口数量约42亿。
克里斯托弗·哥伦布(西班牙语:Cristóbal Colón;意大利语:Cristoforo Colombo,1452年9月22日—1506年5月20日),意大利探险家、殖民者、航海家,大航海时代的主要人物之一。出生于中世纪的热那亚共和国(今意大利西北部)。
汉明帝永平五年(62年),班超举家迁往雒阳(今河南洛阳),日常以替人抄书维持家庭生活。永平十六年(73年),班超以假司马的身份跟随奉车都尉窦固前往边塞,后跟随从事郭恂前往西域。他们辗转停留在西域三十多年,联络各国以孤立匈奴,对国家的巩固统一做出突出贡献。永宁七年(95年),班超被朝廷封为定远侯,后人

提比略·恺撒·奥古斯都罗马帝国第二位皇帝提比略,全名提比略·恺撒·(神君奥古斯都之子·)奥古斯都(拉丁语:Tiberius Caesar Divi Augusti filius Augustus,公元前42年11月16日—公元37年3月16日),原名提比略·克劳狄乌斯·尼禄或提比略·尤里乌斯·恺撒(Tiberius Claudius Nero或Tiberius Julius Caesar),中文又

尚可名片
这家伙太懒了,什么都没写!
作者
